ESTRUCTURA DE LA MEMBRANA
|
|
|
- Guillermo Roldán Lara
- hace 8 años
- Vistas:
Transcripción
1 ESTRUCTURA DE LA MEMBRANA Una célula viva es un sistema de moléculas que se autorreproduce dentro de un recipiente. El recipiente es la membrana plasmática una película lipídica tan delgada y transparente que no puede ser visualizada directamente al microscopio óptico. Su construcción es muy sencilla, basada en una lámina de moléculas lipídicas de aproximadamente 5 nm es decir, de unos 50 átomos de grosor. Sin embargo, sus propiedades son diferentes a las de cualquier otra lámina de un material que nos sea familiar del mundo que nos rodea. Es mucho más que una barrera que impide que el contenido celular se escape y se mezcle con el medio circundante (Figura 11 1A). Los nutrientes han de cruzar la membrana hacia el interior de la célula para que la célula pueda sobrevivir y crecer, y los productos de desecho han de cruzarla hacia el exterior. Así pues, en la membrana hay canales y bombas altamente selectivas, formadas por moléculas proteicas, que permiten que determinadas sustancias sean importadas hacia el interior celular y que otras sustancias sean exportadas hacia el exterior. Además, otras moléculas proteicas actúan de sensores que permiten a la célula responder a los cambios de su entorno. También son destacables las propiedades mecánicas de la membrana. Cuando la célula crece o cambia de forma, también lo hace la membrana plasmática: se alarga, incorporando más membrana sin perder su continuidad, y puede deformarse sin romperse (véase Figura 11-2). Si se agujerea, nunca se colapsa como un globo ni se rompe; al contrario, rápidamente se vuelve a soldar. Figura 11.1 Las membranas celulares como barreras Las membranas celulares entre dos compartimientos entre el interior y el exterior de la célula (A) o entre dos compartimientos intracelulares (B). En cada caso la membrana impide que las moléculas de ambos compartimientos se mezclen 1

2 Figura 11.2 Algunas de las funciones de la membrana plasmática Las bacterias más sencillas tienen una sola membrana la membrana plasmática. En cambio, las células eucariotas tienen, además, una profusión de membranas internas (endomembranas) que envuelven los compartimientos intracelulares. Estas membranas están construidas siguiendo los mismos principios que la membrana plasmática, y también actúan de barrera selectiva entre los espacios que contienen distintas colecciones de moléculas (Figura 11 1B). Así pues, las membranas del retículo endoplásmico, del complejo de Golgi, de las mitocondrias y de los otros orgánulos (Figura 11 3) mantienen las características diferenciales de composición y función entre estos orgánulos. Estas membranas internas actúan mucho más que como barrera: sutiles diferencias entre ellas, especialmente en cuanto a las moléculas proteicas que las forman, son las principales responsables de otorgar a cada orgánulo su carácter diferencial. 2
.")
3 Figura 11.3 Las membranas forman los distintos compartimientos de las células eucariotas En la figura se muestran los distintos orgánulos rodeados de membrana de una típica célula animal. Obsérvese que el núcleo y las mitocondrias están rodeados por dos membranas. Todas las membranas celulares están formadas por lípidos y proteínas y tienen una estructura básica común (Figura 11 4). El componente lipídico está formado por muchos millones de moléculas de lípidos ordenados en dos láminas íntimamente superpuestas, formando una bicapa lipídica (véase Figura 11 4B y C). La bicapa lipídica proporciona la estructura básica y actúa de barrera permeable. Las moléculas proteicas intervienen en el resto de funciones de la membrana y otorgan a las distintas membranas sus características individuales. En este capítulo discutiremos en primer lugar los lípidos de la membrana y luego las proteínas. 3

4 Figura 11.4 Tres perspectivas de la membrana celular (A) Electromicrografía de una membrana plasmática de un eritrocito humano en sección transversal (B) y (C) Dibujos esquemáticos que muestran en dos y tres dimensiones la membrana celular 4
 y (C) Dibujos")
5 La bicapa lipídica La bicapa lipídica se ha establecido como la base universal de la estructura de la membrana celular. Sus propiedades son las responsables de las propiedades generales de las membranas celulares. Empezaremos esta sección considerando de qué forma la estructura de la bicapa es una consecuencia del comportamiento de las moléculas lipídicas en un medio acuoso. Los lípidos de las membranas forman bicapas en el agua Los lípidos de las membranas celulares combinan dos propiedades muy distintas en una misma molécula: tienen una cabeza hidrofílica ("que se siente atraída por el agua") y una o dos colas hidrocarbonadas hidrofóbicas ("que rehúyen el agua") (Figura 11 5). Los lípidos de membrana más abundantes son los fosfolípidos, en los cuales el grupo del extremo hidrofílico se encuentra unido al resto de la molécula por medio de un grupo fosfato. En la mayoría de membranas celulares el tipo más común de fosfolípido es la fosfatidilcolina, que tiene una pequeña molécula de colina unida a un fosfato como cabeza hidrofílica y dos largas cadenas hidrocarbonadas como colas hidrofóbicas (Figura 11 6). Figura 11.5 Dibujo esquemático de una típica molécula lipídica de la membrana 5

6 Figura 11.6 Molécula de fosfatidilcolina La fosfatidilcolina, representada ( A ) de forma esquemática (B) como fórmula química (C) como modelo espacial compacto (D) como símbolo Ese fosfolípido tiene cinco partes: la cabeza hidrofílica, la colina unida por el fosfato al glicerol; que está unido a las dos cadenas hidrocarbonadas que forman la cola hidrofóbicas. Las dos cadenas son sintetizadas como ácidos grasos cadenas hidrocarbonadas con un grupo COOH en un extremo- que están unidas al glicerol por sus grupos COOH. Cuando hay un doble enlace entre dos carbonos se produce un doblegamiento de las cadenas hidrocarbonadas; la curvatura debida al doble enlace está exagerada en los dibujos. El término fosfatidil que forma parte del nombre de los fosfolípidos se refiere a la parte fosfato glicerol ácido graso de la molécula. 6

7 Todas las moléculas con propiedades hidrofílicas e hidrofóbicas reciben el nombre de anfipáticas. Esta característica también la presentan otros tipos de lípidos de membrana los esteroles (como el colesterol en las membranas celulares animales) y los glucolípidos, que tienen azúcares en su cabeza hidrofilica (Figura 11 7) y juega un papel muy importante en el ensamblaje de los lípidos que forman la bicapa. Figura 11.7 Tres tipos distintos de moléculas lipídicas de membrana Todas son antipáticas, con una cabeza hidrofílica y una o dos colas hidrofóbicas. La cabeza (sombreada en rosa) es una serina fosfato en la fosfatidilserina, un grupo OH en el colesterol, y un azúcar (galactosa) en el galactocerebrósido Como hemos discutido en el Capítulo 2, las moléculas hidrofílicas se disuelven fácilmente en el agua puesto que disponen de átomos cargados o grupos polares (es decir, grupos con una 7

8 distribución desigual de cargas positivas y negativas) que pueden formar enlaces hidrostáticos o enlaces de hidrógeno con las moléculas de agua, las cuales también son polares (Figura 11 8). Por el contrario, las moléculas hidrofóbicas son insolubles en el agua puesto que todos, o casi todos, sus átomos son apolares o sin carga y, por tanto, no pueden formar enlaces con las moléculas de agua. En lugar de disolverse, fuerzan a las moléculas de agua adyacentes a reorganizarse adoptando una estructura parecida a una jaula alrededor de la molécula hidrofóbica (Figura 11 9). Esta estructura es mucho más ordenada que el agua que la envuelve, por lo que su formación necesita energía. Sin embargo, el coste energético es mínimo si las moléculas hidrofóbicas se agrupan de manera que quede afectado el número mínimo de moléculas de agua. Así pues, las moléculas puramente hidrofóbicas, como las grasas de las células adiposas animales y los aceites de las semillas de las plantas (Figura 11 10), coalescen en una única gran gota cuando se dispersan en el agua. Figura 11.8 Una molécula hidrofílica interactuando con moléculas de agua La acetona es una molécula polar, por los que puede formar interacciones favorables con las moléculas de agua, las cuales también son polares. Así pues la acetona se disuelve fácilmente en agua. δ- indica una carga parcial negativa y δ+ una carga parcial positiva. En la figura los átomos polares se han coloreados en rosa y azul y los grupos no polares están en gris. Figura 11.9 Una molécula hidrofóbica en el agua La molécula de 2 metil propano es completamente Hidrofóbica, por lo que no puede formar interacciones favorables con el agua y fuerza a las moléculas de agua adyacentes a reorganizarse en una estructura en forma de caja a su alrededor. 8

9 Figura Comparación entre una molécula grasa (triacilglicerol) y una molécula de fosfolípido (fosfatidiletanolamina) Las zonas hidrofóbicas están sombreadas en gris, y las hidrofílicas en rosa. La molécula de grasa es totalmente hidrofóbica mientras que la molécula de fosfolípido es antipática. (En la figura, la tercera cola del triacilglicerol está situada hacia arriba de la molécula para poderla comparar con el fosfolípido, aunque normalmente se encuentra hacia abajo) Las moléculas antipáticas como los fosfolípidos están sujetas a dos fuerzas conflictivas: la cabeza hidrofílica es atraída por el agua, mientras que las colas hidrofóbicas rehuyen el agua y tienden a agregarse a otras moléculas hidrofóbicas. Este conflicto se resuelve maravillosamente con la formación de una bicapa lipídica una disposición que satisface ambas partes y es lo más favorable desde el punto de vista energético. Las cabezas hidrofílicas quedan expuestas al agua mientras que todas las colas hidrofóbicas quedan alejadas del medio acuoso, unas junto a otras en el interior del sándwich (Figura 11 11). 9

10 Figura Visión transversal de una bicapa de fosfolípidos. (A) Simulación por ordenador que muestra las moléculas de fosfolípido (Colas rojas y cabezas amarillas) y las moléculas de agua que las rodean (azul). (B) Dibujo esquemático de una bicapa de fosfolípidos en el agua. (Adaptado de Science 262: (1993), cortesía de R. Venable y R. Pastor.) Las mismas fuerzas que conducen a las moléculas anfipáticas a formar una bicapa confieren a é sta bicapa su capacidad autoselladora. Cualquier rotura de la bicapa dará lugar a un extremo libre en el agua, y puesto que esto es energéticamente desfavorable, las moléculas de la bicapa se reordenarán espontáneamente, con lo cual la bicapa será reparada, restableciéndose de nuevo una capa continua. Si la fisura es muy grande, la capa puede romperse en vesículas aisladas. En estos casos, el principio predominante es que los extremos 10
 Las mismas fuerzas que conducen a las moléculas anfipáticas a formar una bicapa confieren a é sta bicapa su capacidad autoselladora.")
11 libres sean rápidamente eliminados. La prohibición de que existan extremos libres tiene una consecuencia muy importante: sólo existe un sistema para que una lámina finita pueda evitar tener extremos libres, que consiste en cerrarse sobre sí misma y originar un espacio cerrado (Figura 11 12). Así pues, las moléculas antipáticas como los fosfolípidos se ensamblan necesariamente formando recipientes que se autosellan definiendo así compartimientos cerrados. Este comportamiento tan importante, fundamental para la creación de una célula viva, es en esencia sencillamente el resultado del hecho de que cada molécula es hidrofílica en un extremo e hidrofóbica en el otro. Figura Las bicapas de fosfolípidos se doblan espontáneamente formando comportamientos cerrados. La estructura cerrada es estable puesto que evita la exposición de las colas hidrocarbonadas hidrófóbicas al agua, hecho energéticamente desfavorable. 11

12 La bicapa lipídica es un fluido bidimensional El ambiente acuoso dentro y fuera de la célula impide a los lípidos de la membrana escaparse de a bicapa, pero nada impide que estas moléculas se desplacen y cambien de posición las unas respecto a las otras dentro del plano de la bicapa. Por lo tanto, la membrana se comporta como un fluido bidimensional, lo cual es crucial para sus funciones. Esta propiedad es distinta de la de flexibilidad, que es la capacidad de la membrana para doblegarse. La flexibilidad de la membrana también es importante, y establece un límite menor de aproximadamente 25nm para el tamaño de la vesícula que las membranas celulares pueden formar. La fluidez de las bicapas lipídicas puede estudiarse utilizando bicapas lipídicas sintéticas, que pueden generarse fácilmente de manera artificial mediante la agregación espontánea de moléculas lipídicas anfipáticas en el agua. En estudios experimentales se suelen utilizar dos tipos de estas bicapas sintéticas. Al añadir fosfolípidos puros al agua se forman vesículas esféricas cerradas, llamadas liposomas; son de tamaño variable, desde 25nm hasta 1mm de diámetro (Figura 11 13). Alternativamente, se pueden formar bicapas de fosfolípidos planas a través de un agujero situado en una separación entre dos compartimientos acuosos (Figura 11 14). Figura Liposomas Electrónmicrografía de vesículas de fosfolípidos (liposomas) que muestra la estructura de la bicapa de la membrana. Esquema de un pequeño liposomas esférico visto en sección transversal. 12

13 Figura Representación de una sección transversal de una bicapa lipídica sintética Esta bicapa plana se forma a través de un pequeño agujero (aproximadamente de 1mm de diámetro) en una pared que separa dos compartimientos acuosos. Para formar la bicapa en el agujero, el agujero se sumerge en una solución acuosa y la solución de fosfolípidos (en disolvente no acuoso) se pinta a través del agujero con un pincel. Estas bicapas artificiales sencillas permiten medidas delicadas del desplazamiento de las moléculas lipídicas, y revelan que algunos desplazamientos son raros mientras que otros son rápidos y frecuentes. Así pues, en las bicapas lipídicas sintéticas, las moléculas de fosfolípidos raramente saltan de un lado de la monocapa (una mitad de la bicapa) al otro; sin proteínas que lo faciliten y en condiciones similares a las de la célula, se estima que este proceso, denominado "flip-flop", se produce menos de una vez al mes en cualquier molécula lipídica. Por otro lado, debido a movimientos térmicos, las moléculas lipídicas giran muy rápidamente sobre su eje longitudinal y constantemente intercambian su lugar con el de las moléculas vecinas dentro de cada monocapa (Figura 11 15). Este intercambio da lugar a una rápida difusión lateral; por ejemplo, una molécula lipídica en una bicapa artificial puede difundir una longitud igual a la de una gran célula bacteriana (~2µm) en aproximadamente un segundo. Si la temperatura disminuye, la caída de la energía térmica hace disminuir la velocidad del movimiento de los lípidos, con lo que la membrana es menos fluida. 13
 se pinta a través del agujero con un pincel.")
14 Se han obtenido resultados parecidos con membranas celulares aisladas y con células enteras, lo cual indica que la bicapa lipídica de una membrana celular también se comporta como un fluido bidimensional en el que las moléculas constituyentes son libres de desplazarse lateralmente; como en las bicapas sintéticas, en las células las moléculas individuales de fosfolípido normalmente se hallan restringidas a su propia monocapa y no sufren espontáneamente el proceso de "flip-flop" (véase Figura 11 15). Figura Movilidad de los fosfolípidos Tipos de movimientos posibles de las moléculas de fosfolípidos en una bicapa lipídica. La fluidez de una bicapa lipídica depende de su composición El grado de fluidez de una membrana celular la facilidad con la que sus moléculas lipídicas se desplazan en el plano de la bicapa es importante para su función y se ha de mantener dentro de unos ciertos límites. La fluidez de una bicapa lipídica a una temperatura determinada depende de su composición de fosfolípidos y, especialmente, de la naturaleza de las colas 14

15 hidrocarbonadas: cuanto más regular y más compacto sea el empaquetamiento de estas colas, más viscosa y menos fluida será la bicapa. Dos propiedades de las colas hidrocarbonadas afectan su empaquetamiento en la bicapa su longitud y su insaturación (es decir, el número de dobles enlaces que contienen). Los extremos hidrocarbonados de las moléculas de fosfolípidos varían entre 14 y 24 átomos de carbono de longitud, siendo lo más habitual átomos. Una longitud menor de la cadena reduce la tendencia de las colas hidrocarbonadas para interaccionar entre sí, de forma que aumenta la fluidez de la bicapa. Generalmente una de las dos colas hidrocarbonadas de cada molécula de fosfolípido tiene uno o más dobles enlaces entre átomos de carbono adyacentes (véase Figura 11 6). Por lo tanto, no tiene el número máximo de átomos de carbono que en principio podrían unirse a su esqueleto carbonado, por lo que se dice que está insaturada respecto a los hidrógenos. Generalmente la otra cola de ácido graso no tiene dobles enlaces, es decir, presenta el complemento completo de átomos de hidrógeno, con lo cual está saturada. Cada doble enlace de una cola insaturada genera un pequeño remolino en la cola hidrocarbonada (véase Figura 11 6) que dificulta el empaquetamiento de las colas. Por esta razón, las bicapas lipídicas que tienen grandes cantidades de colas hidrocarbonadas insaturadas son mucho más fluidas que las que tienen proporciones menores. Las bacterias y las levaduras, que han de adaptarse a temperaturas muy variables, ajustan constantemente tanto la longitud como la insaturación de las colas hidrocarbonadas de los fosfolípidos de su bicapa para mantener relativamente constante la fluidez de la membrana: por ejemplo, a temperaturas altas la célula fabrica lípidos de membrana con colas más largas y que contienen muy pocos dobles enlaces. Se utiliza un truco parecido en la fabricación de la margarina a partir de aceites vegetales. Las grasas producidas por las plantas son generalmente insaturadas y, por tanto, líquidas a temperatura ambiente, a diferencia de las grasas animales tales como la mantequilla o el sebo, que están generalmente saturadas y, por lo tanto, a esta misma temperatura son sólidas. La margarina se fabrica con aceites vegetales a los cuales se les han eliminado los dobles enlaces añadiéndoles hidrógeno, con lo que a temperatura ambiente son más sólidos y parecidos a la mantequilla. 15

16 La fluidez de la membrana es importante para la célula por muchas razones. Permite a las proteínas de membrana difundir rápidamente en el plano de la bicapa y que interaccionen unas con otras, lo cual es muy importante, por ejemplo, en los procesos de señalización celular (se discute en el Capítulo 15). Proporciona una manera sencilla de distribuir los lípidos y las proteínas, mediante difusión desde los lugares donde han sido insertados en la bicapa después de su síntesis, hasta otras regiones de la célula. Permite a las membranas fusionarse unas con otras y mezclar sus moléculas, y permite que las moléculas de la membrana se distribuyan de forma equitativa entre las dos células hijas durante la división celular. Es difícil imaginarse cómo podría una célula vivir, crecer y reproducirse si sus membranas no fueran fluidas. En las células animales, la fluidez de la membrana está modulada por la presencia del esterol colesterol, ausente en plantas, levaduras y bacterias. Estas cortas y rígidas moléculas se encuentran en grandes cantidades en la membrana plasmática, donde rellenan los espacios existentes entre las moléculas vecinas de fosfolípidos provocados por los remolinos de sus extremos hidrocarbonados insaturados (Figura11 16). De esta forma el colesterol endurece la bicapa y la hace menos fluida y menos permeable. 16

17 Figura Esquemas superiores Estructura del colesterol representada mediante: (A) su fórmula (B) un esquema (C) un modelo compacto espacial. Esquema inferior: Papel del colesterol en las membranas celulares El colesterol en una bicapa lipídica. Dibujo esquemático de una molécula de colesterol interactuando con 2 moléculas de fosfolípido en una de las monocapas de la bicapa lipídica. 17

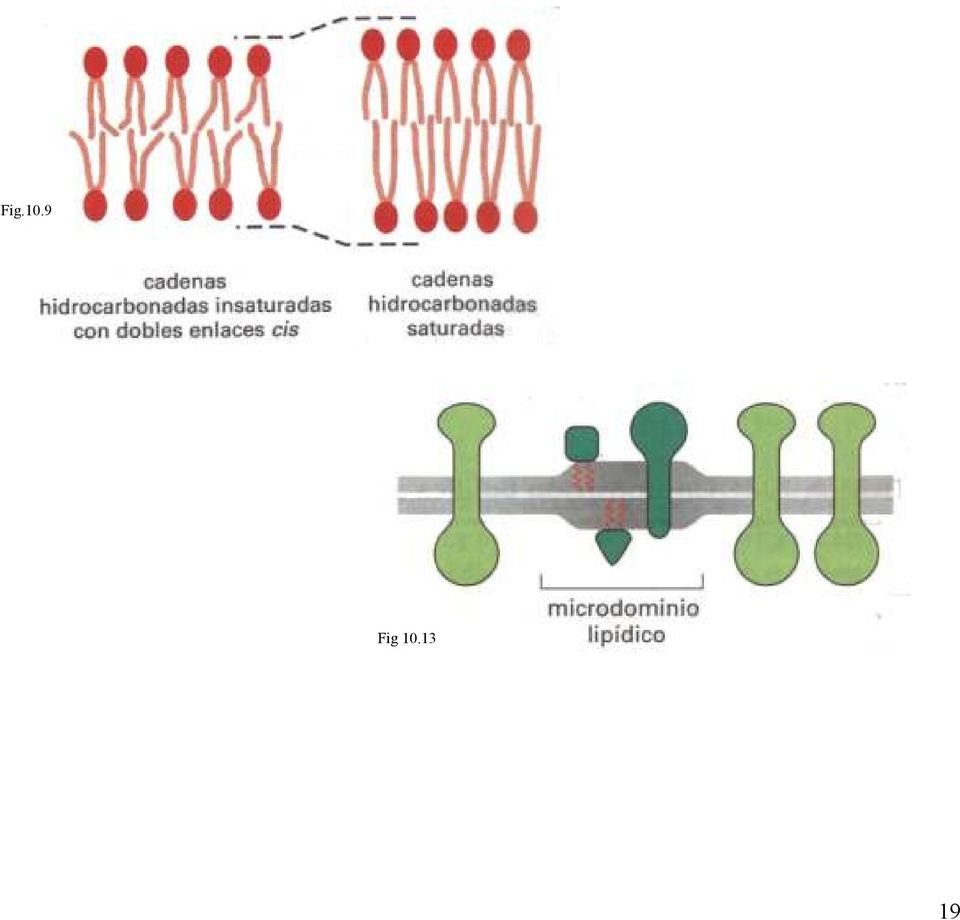
18 ACTUALIZACION 1 La membrana plasmática contiene microdominios lipídicos enriquecidos en esfingolípidos, colesterol y algunas proteínas de membrana. La mayoría de los diferentes tipos de moléculas lipídicas de las membranas celulares están distribuidas al azar mezcladas unas con otras en la monocapa en la que residen. Las fuerzas de atracción de van der Waals entre los ácidos grasos adyacentes no son tan selectivas como para mantener unidos grupos de moléculas de este tipo. Sin embargo, para algunas moléculas lipídicas, como los esfingolípidos que suelen presentar cadenas hidrocarbonadas saturadas largas, las fuerzas de atracción podrían ser lo suficientemente fuertes como para mantener unidas temporalmente las moléculas adyacentes formando pequeños microdominios. Estos microdominios lipídicos pueden considerarse una transición de fase temporal en la bicapa lipídica en la que los esfingolípidos se concentran. Parece que la membrana plasmática de las células animales contiene muchos pequeños microdominios lipídicos (de aproximadamente 70nm de diámetro), ricos tanto en esfingolípidos como en colesterol. Las cadenas hidrocarbonadas de los lípidos concentrados son más largas y rectas que las de la mayoría de los lípidos de membrana por lo que los microdominios son más gruesos que el resto de la bicapa (fig. 10.9) y pueden acomodar con mayor facilidad ciertas proteínas de membrana que tienden a acumularse en ellos (fig ). Así, parece que los microdominios lipídicos favorecen la organización de estas proteínas concentrándolas para su transporte en pequeñas vesículas o permitiendo que puedan actuar unidas, como cuando transducen señales extracelulares en señales intracelulares. Casi siempre las moléculas lipídicas de una monocapa de la bicapa se desplazan independientemente de las de la otra monocapa. Sin embargo en los microdominios lipídicos las largas cadenas hidrocarbonadas de los esfingolípidos de una monocapa interaccionan con las de otra monocapa, de forma que en estos microdominios lipídicos las dos monocapas se comunican a través de sus colas lipídicas. Tomado de ALBERTS B.Biología molecular de la célula (4ª edición 2004) Editorial Omega 18

19 Fig.10.9 Fig
20 ACTUALIZACION 2 Las gotas de lípido están rodeadas por una monocapa de fosfolípidos La mayoría de las células almacena un exceso de lípidos como gotas, a partir de las cuales se obtienen bloques constructores para la síntesis de la membrana o como fuente de alimento. Los adipocitos son células especializadas en el almacenamiento de lípidos. Tienen un alto contenido de gotas lipídicas de las cuales los ácidos grasos pueden ser liberados por demanda y exportados a otras células a través del torrente sanguíneo. Las gotas de lípidos almacenan lípidos neutros tales como triacilglicéridos y ésteres del colesterol, los cuales son sintetizados por enzimas a partir de ácidos grasos y colesterol en la membrana del retículo endoplásmico. Como estos lípidos no contienen grupos hidrofílicos, son moléculas hidrofóbicas, se agregan en gotas tridimensionales más que en bicapas. Las gotas de lípidos son organelos únicos porque están rodeados por una monocapa de fosfolípidos, la cual contiene una gran variedad de proteínas. Algunas de éstas son enzimas involucradas en el metabolismo de los lípidos, pero de la mayoría se desconoce su función. Las gotas de lípidos se forman rápidamente cuando las células son expuestas a una alta concentración de ácidos grasos. Se forman a partir de discretas regiones de la membrana del retículo endoplásmico donde están concentradas varias de las enzimas relacionadas con el metabolismo lipídico. En la figura se muestra un modelo de cómo las gotas lipídicas podrían formarse y adquirir la monocapa que las rodea. Tomado de ALBERTS B.Biología molecular de la célula (5ª edición 2004) Editorial Omega 20

21 21
22 La bicapa lipídica es asimétrica Generalmente las membranas celulares son asimétricas, presentando un aspecto muy distinto en el interior de la célula o del orgánulo que en el exterior. Frecuentemente las dos mitades de una bicapa presentan diferencias importantes en cuanto a su composición de fosfolípidos y de glucolípidos (Figura 11 17). Además, las proteínas están embebidas en la bicapa en una orientación específica, muy importante para su función. Figura La distribución asimétrica de los fosfolípidos y de los glucolípidos en la bicapa lipídica de una membrana plasmática La Se asimetría muestran en lipídica distintos empieza colores cinco en tipos el de momento moléculas de fosfolípidos su síntesis. (marcados En en las letra células, roja), En las azul, nuevas los moléculas glucolípidos, de con fosfolípido grupos de son cabeza sintetizadas hexagonales por que enzimas representan unidas los azúcares. a la membrana, Todas las moléculas que utilizan de como glucolípidos se encuentran en la monocapa externa, mientras que el colesterol se distribuye por igual en ambas sustratos ácidos grasos disponibles en una de las dos mitades de la bicapa esto es, en una monocapas. monocapa y liberan el nuevo fosfolípido en la misma monocapa. Para permitir que la 22
23 membrana entera crezca, una proporción de las moléculas lipídicas se ha de trasladar a la monocapa opuesta. Esta transferencia está catalizada por unas enzimas llamadas flipasas (Figura 11 18). Se ha sugerido que las flipasas pueden transferir selectivamente moléculas específicas de fosfolípido, con lo cual en las dos mitades de la bicapa habría distintos tipos de flipasas. Figura Papel de las flipasas en las síntesis de las bicapas lipídicas Todas las moléculas de fosfolípido de nueva síntesis se añaden a una de las caras de la bicapa, pero las flipasas transfieren algunas de ellas a la monocapa opuesta, con lo cual la bicapa se alarga La inserción en una sola cara y la presencia de flipasas selectivas no son los únicos sistemas de producir bicapas lipídicas asimétricas. En el caso de los glucolípidos, el tipo de moléculas lipídicas que presentan la distribución asimétrica más consistente y sorprendente, actúa un mecanismo distinto. Para explicar su distribución es necesario conocer con más detalle la ruta de formación de membrana en las células eucariotas. 23
24 La asimetría lipídica se genera dentro de la célula En las células eucariotas la síntesis de la membrana se produce en un compartimiento intracelular el retículo endoplásmico, o ER (de "endoplasmic reticulum", discutido con más detalle en el Capítulo 14). La nueva membrana, una vez ensamblada, es exportada hasta las otras membranas de la célula mediante un ciclo de gemación y fusión: del retículo endoplásmico se desprenden pequeñas porciones de membrana, formando pequeñas vesículas, las cuales se incorporan mediante fusión a otras membranas. Durante este proceso de transporte vesicular se conserva la orientación de la bicapa respecto al citosol, por lo que todas las membranas celulares, tanto la membrana plasmática externa como las intracelulares que rodean los orgánulos, tienen caras "internas" y "externas" distintas: la interna, o citosólica, limita con el citosol, mientras que la externa, o no citosólica, está expuesta al exterior celular o al espacio interior de un orgánulo (Figura 11 19). Los glucolípidos están localizados principalmente en la membrana plasmática, y se encuentran únicamente en la mitad no citosólica de la bicapa. Por lo tanto, sus grupos de azúcar están expuestos al exterior celular (véase Figura 11 17), donde forman parte de la cubierta protectora de hidratos de carbono que rodea la mayoría de las células animales. Las moléculas de glucolípido adquieren sus grupos azúcar en el complejo de Golgi (se discute en el Capítulo 14). Las enzimas que añaden estos grupos se encuentran localizadas en el interior del complejo de Golgi, con lo cual las moléculas de azúcar se añaden a las moléculas lipídicas en la mitad no citosólica de la bicapa lipídica. Cuando se ha formado una molécula de glucolípido, permanece atrapada en esta monocapa, puesto que allí no hay flipasas que puedan transferirla a la monocapa citosólica. Así pues, cuando finalmente llega a la membrana plasmática, la molécula de glucolípido está situada de forma que su azúcar está colocado hacia el exterior de la célula (véase Figura 11 19). 24
25 Figura Aproximación y fusión de una vesícula de membrana Una vesícula de membrana aproximándose a un orgánulo rodeado de membrana y fusionándose con su membrana plasmática. Obsérvese cómo durante el proceso de aproximación y fusión se mantiene la orientación de la membrana, de forma que la superficie citosólica sigue siendo la superficie citosólica. Otras moléculas lipídicas presentan tipos distintos de distribución asimétrica, relacionados con otras funciones. Por ejemplo, los fosfolípidos del tipo inositol son componentes minoritarios de la membrana plasmática, pero juegan un papel especial en la transmisión de señales desde la superficie celular hasta los compartimientos intracelulares que responderán a la señal (se discute en el Capítulo 15). Actúan con posterioridad a que la señal haya sido transmitida a través de la membrana plasmática, por lo que están concentrados en la mitad citosólica de la bicapa lipídica (véase figura 11-17). 25
26 Las bicapas lipídicas son impermeables a los solutos y a los iones Hemos visto que una de las funciones más importantes de las membranas celulares es el control del paso de moléculas a través de la bicapa lipídica. En este aspecto el interior hidrofóbico de la bicapa lipídica juega un papel muy importante, puesto que constituye una barrera al paso de la mayoría de moléculas hidrofílicas. Estas moléculas rehúyen entrar en un ambiente graso, como lo hacen las moléculas hidrofóbicas a entrar en el agua. Esta función de barrera de las bicapas lipídicas se puede poner de manifiesto en bicapas sintéticas del tipo que se ilustra en la Figura Con tiempo suficiente, prácticamente cualquier molécula acabará difundiendo a través de una bicapa lipídica. Sin embargo, la velocidad a la que se produce esta difusión varía enormemente, dependiendo en parte del tamaño de la molécula y, principalmente, de sus propiedades de solubilidad. En general, cuanto menor es la molécula y cuanto más soluble es en aceite (es decir, cuanto más hidrofóbica o no polar sea) más rápidamente difunde a través de una bicapa. Así pues: 1. Moléculas pequeñas no polares, tales como el oxígeno molecular (O2, de 32 daltons de peso molecular) y el dióxido de carbono (44 daltons), se disuelven fácilmente en las bicapas lipídicas y por lo tanto difunden con rapidez a través de ellas; de hecho, las células necesitan ser permeables a los gases para los procesos de respiración celular, los cuales se tratan en el Capítulo Moléculas polares no cargadas (moléculas con una distribución desigual de la carga eléctrica) también difunden rápidamente a través de una bicapa, si su tamaño es suficientemente reducido. Por ejemplo, el agua (18 daltons) y el metanol (46 daltons) atraviesan rápidamente una bicapa; el glicerol (92 daltons) lo hace con menor rapidez, y la glucosa (180 daltons) prácticamente no la atraviesa (Figura 11-20). 3. Por el contrario, las bicapas lipídicas son altamente impermeables a todos los iones y moléculas cargadas, por muy pequeñas que sean. Su carga y el elevado grado de 26
27 atracción hacia las moléculas de agua les impiden penetrar en la fase hidrocarbonada de la bicapa. Así, las bicapas artificiales son mil millones (10 9 ) de veces más permeables al agua que a los iones, incluso a los de reducido tamaño como el Na + o el K +. Figura Permeabilidad relativa de una bicapa lipídica artificial a diferentes tipos de moléculas Cuanto más pequeña sea la molécula y, lo que es más importante, cuanto menor sea el número de interacciones con el agua (es decir, cuanto menos polar sea), más rápidamente difundirá a través de una bicapa. Obsérvese que muchas de las moléculas que utilizan las células como nutrientes son demasiado grandes y polares para cruzar a través de una bicapa pura. 27
28 Por lo tanto, las membranas celulares permiten el paso, por simple difusión, del agua y de las moléculas no polares. Sin embargo, para que las células puedan captar nutrientes y eliminar los productos de deshecho, las membranas también han de permitir el paso de muchas otras moléculas, tales como iones, azúcares, aminoácidos, nucleótidos y muchos metabolitos celulares. Estas moléculas cruzan las bicapas lipídicas con gran lentitud por difusión simple, por lo que son necesarias proteínas de transporte especializadas para transportarlas de manera eficiente a través de las membranas celulares. Estas proteínas transportadoras de membrana son objeto de estudio en el Capítulo 12. Sin embargo, antes de entrar a estudiar este punto, vamos a considerar algunos principios generales sobre cómo están asociadas las proteínas con la bicapa lipídica en las membranas celulares Proteínas de membrana La bicapa lipídica es la estructura básica de todas las membranas biológicas y actúa de barrera de permeabilidad, pero la mayoría de las funciones específicas de la membrana las desempeñan las proteínas de membrana. En las células animales, las proteínas constituyen cerca del 50% de la masa total de la mayoría de membranas plasmáticas, mientras que la otra mitad de la masa son lípidos y una cantidad relativamente pequeña de carbohidratos. Sin embargo, las moléculas lipídicas son mucho más pequeñas que las moléculas de proteína, por lo que por cada molécula de proteína hay alrededor de 50 moléculas de lípidos (véase Figura 11 4). 28
29 Figura Algunas funciones de las proteínas de la membrana plasmática Las proteínas de membrana realizan muchas otras funciones, además de transportar nutrientes, metabolitos, o iones a través de la bicapa lipídica. Algunas de ellas unen macromoléculas a la membrana, a uno u otro lado. Otras actúan como receptores que detectan señales químicas en el ambiente celular y las transmiten al interior de la célula; otras, como las enzimas, catalizan reacciones específicas (Figura 11-21; Tabla11-1). Cada tipo de membrana presenta un juego diferente de proteínas, lo cual refleja las funciones particulares de una membrana determinada. En esta parte del capítulo discutiremos la estructura de las proteínas de membrana e ilustraremos las distintas maneras en que se asocian con la bicapa lipídica. 29
30 Tabla 11 1 Algunos ejemplos de proteínas de membrana plasmática y sus funciones Clase funcional Ejemplo de proteínas Función específica Transportadores bomba de Na + bombea en forma activa Na+ hacia el exterior y K + hacia el interior de la célula Conectores Receptores Enzimas integrinas receptor del factor de crecimiento derivado de las plaquetas (PDGF) adenilato ciclasa unen los filamentos intracelulares de actina con las proteínas de la matriz extracelular une el PDGF extracelular y, como consecuencia, genera una señal intracelular que provoca que la célula crezca y se divida cataliza la producción de AMP cíclico intracelular como respuesta a señales intracelulares Las proteínas de membrana pueden estar asociadas a la bicapa lipídica de varias maneras Las proteínas se pueden asociar a la bicapa lipídica de una membrana celular de tres maneras principales (Figura 11-22). 1. Muchas proteínas de membrana atraviesan la bicapa lipídica de forma que parte de su masa se sitúa a cada lado de la membrana (Figura 11 22A). Estas proteínas transmembrana, al igual que sus vecinas lipídicas, tienen regiones hidrofílicas e hidrofóbicas. Las regiones hidrofóbicas se sitúan en el interior de la membrana en contacto con las colas hidrofóbicas de las moléculas lipídicas. Sus regiones hidrofílicas se hallan expuestas al medio acuoso de ambos lados de la membrana. 30
31 2. Otras proteínas de membrana se hallan localizadas completamente en el exterior de la bicapa, unidas a la bicapa únicamente por medio de una o más uniones covalentes con grupos de lípidos (11 22B). 3. Otras proteínas están unidas de manera indirecta a un lado u otro de la membrana sólo mediante interacciones con otras proteínas de membrana (Figura 11 22C). Todas las proteínas de membrana presentan una orientación característica: por ejemplo, una proteína transmembrana siempre tiene la misma región proteica en contacto con el citosol. La orientación es una consecuencia de la manera por la que se ha sintetizado la proteína, como se discute en el Capítulo 14. Figura Distintas formas de asociación de las proteínas de membrana con la bicapa lipídica (A) Las proteínas transmembrana pueden atravesar la bicapa como hélices α sencillas, como hélices α múltiples, como láminas β cerradas (barril β). (B) Otras proteínas de membrana están unidas a la bicapa solamente mediante unión covalente a las moléculas de lípidos (líneas rojas en zig zag). (C) Finalmente, muchas proteínas de membrana están unidas a la membrana mediante interacciones no covalentes relativamente débiles con otras proteínas de membrana. 31
32 ACTUALIZACION 3 Tomado de ALBERTS B.Biología molecular de la célula (4ª edición 2004) Editorial Omega Varios sistemas de asociación de las proteínas con la bicapa lipídica Proteínas transmembrana que atraviesan la bicapa: (1) como una α-hélice única (de paso único). (2) en forma de múltiples α-hélices (de paso múltiple) Algunas de estas proteínas de paso único o de paso múltiple están unidas covalentemente a cadenas de ácidos grasos insertados en la monocapa citoplasmática (3) en forma de hebra β enrollada (un barrilβ). Otras proteínas están expuestas sólo a un lado de la membrana: (4) algunas de ellas están unidas a la superficie citosólica mediante una α hélice anfipática que se une a la monocapa citosólica a través de la superficie hidrofóbica de la hélice. (5) otras se encuentran unidas a la BL únicamente a través de su unión covalente a una cadena de lípido (a una cadena de ácido graso o a un grupo prenil) de la monocapa citosólica (6) a través de una unión mediante un oligosacárido, a un fosfatidilinositol de la monocapa no citosólica. Proteínas periféricas (7) y (8) unidas a través de interacciones no covalentes con otras proteínas de membrana 32
33 Las proteínas que están unidas directamente a la membrana -ya sean transmembrana o unidas a lípidos solamente se pueden disociar de las membranas rompiendo la bicapa lipídica con detergentes, como se discute más adelante. Estas proteínas se conocen como proteínas integrales de membrana. Al resto de proteínas se las conoce como proteínas periféricas de membrana; éstas pueden ser liberadas de la membrana mediante procedimientos de extracción relativamente suaves que interfieren con las interacciones proteicas pero que mantienen intacta la bicapa lipídica. Una cadena polipeptídica habitualmente cruza la bicapa mediante una conformación de hélice α Las zonas de una proteína transmembrana que se localizan fuera de la bicapa lipídica están conectadas unas con otras mediante segmentos especializados de la cadena polipeptídica que atraviesan la membrana. Estos segmentos, que atraviesan el ambiente hidrofóbico del interior de la bicapa lipídica, están formados principalmente por aminoácidos cuyas cadenas laterales son hidrofóbicas. Debido a que estas cadenas laterales no pueden interaccionar con las moléculas de agua, prefieren el ambiente lipídico, en el que no hay agua. Sin embargo, al contrario que las cadenas laterales hidrofóbicas, los enlaces peptídicos que unen los aminoácidos sucesivos de una proteína normalmente son polares, lo cual determina que el esqueleto polipeptídico sea hidrofilico (Figura 11 23). Debido a que en la bicapa no hay agua, todos los enlaces peptídicos de la zona de la cadena que atraviesa la bicapa tienden a formar enlaces de hidrógeno entre ellos. Este tipo de uniones se hace máximo si la cadena polipeptídica que cruza la bicapa forma una hélice α regular; se cree que la mayoría de los segmentos de las cadenas polipeptídicas que atraviesan la membrana se hallan en esta conformación. En este tipo de hélice α, los aminoácidos hidrofóbicos de las cadenas laterales están expuestos al exterior de la hélice, donde entran en contacto con las colas hidrofóbicas de los lípidos, mientras que las distintas partes de] esqueleto polipeptídico forman enlaces de hidrógeno entre sí en el interior de la hélice (Figura 11 24). 33
34 Figura Enlaces peptídicos Los enlaces peptídicos (sombreados en gris) que unen loa aminoácidos adyacentes en una cadena polipeptídica son polares y pt lo tanto hidrofilitos δ- indica una carga parcial negativa y δ+ indica una carga parcial positiva. Figura Un fragmento de una hélice que atraviesa una bicapa lipídica Las cadenas laterales hidrofóbicas de los aminoácidos que forman las hélices α entran en contacto con las colas hidrofóbicas hidrocarbonadas de las moléculas de fosfolípido, mientras que las zonas hidrofílicas del esqueleto polipeptídico forman enlaces de hidrógeno entre ellas, por el interior de la hélice. Se necesitan aproximadamente 20 aminoácidos para atravesar la membrana de esta forma. 34
35 La cadena polipeptídica de muchas proteínas transmembrana cruza la membrana una sola vez (véase Figura 11-22A). Muchas de estas proteínas son receptores de señales extracelulares; su extremo extracelular se une a la molécula señalizadora, mientras que su porción citoplasmática transmite la señal hacia el interior celular (véase Figura 11 21). Otras proteínas transmembrana forman poros acuosos que permiten el paso a través de la membrana de moléculas solubles en agua. Estos poros no los pueden formar las proteínas que tienen una sola hélice α transmembrana, uniformemente hidrofóbica. Son necesarias proteínas transmembrana mucho más complejas, en las que la que la cadena polipeptídica cruza la bicapa varias veces, como hélice α o como un barril cerrado (denominado barril β ) (véase Figura 11 22A). Muchas de estas proteínas tienen una o más regiones transmembrana formadas a partir de hélices α que contienen aminoácidos con cadenas laterales hidrofilicas y con cadenas laterales hidrofóbicas. Las cadenas laterales hidrofóbicas yacen en un lado de la hélice, expuestas a los lípidos de la membrana. Las cadenas laterales hidrofílicas están concentradas en el otro lado, donde forman parte de un poro hidrofílico creado mediante el empaquetamiento de distintas hélices que forman un tubo dentro del ambiente hidrofóbico de la bicapa lipídica (Figura 11 25). En el Capítulo 12 se discute cómo actúan estos poros en el transporte selectivo de pequeñas moléculas solubles en el agua, Figura Formación de un poro hidrofílico transmembrana mediante varias hélices En este ejemplo, cinco hélices α transmembrana forman un canal relleno de agua a través de la bicapa lipídica. Las cadenas laterales de aminoácidos hidrófobicos (verde) en uno de los lados de cada hélice entran en contacto con las colas hidrocarbonadas hidrofóbicas, mientras que las cadena laterales hidrofílicas (rojo) en el lado opuesto de las hélices forman el poro relleno de agua. 35
36 Sin lugar a dudas, la hélice α es la forma más común mediante la cual una cadena polipeptídica cruza la bicapa lipídica, pero algunas proteínas transmembrana pueden cruzarla en estructura de lámina β (discutido en el capítulo 5), curvada formando un cilindro en forma de barril que se abre y cierra, llamado barril β. Como era de esperar, las cadenas laterales de los aminoácidos que están en contacto con el interior del barril, y por lo tanto forman un canal acuoso, son principalmente hidrofilicas, mientras que las cadenas laterales hidrofóbicas están en el exterior, en contacto con el núcleo hidrofóbico de la bicapa lipídica. El ejemplo más representativo de este tipo de proteínas transmembrana son las proteínas llamadas porinas, que forman grandes poros llenos de agua a través de la membrana externa de las mitocondrias y de algunas bacterias, permitiendo el paso de nutrientes y pequeños iones, pero que impiden la entrada de antibióticos y toxinas. A diferencia de las hélices α, los barriles β solamente pueden formar canales anchos, puesto que existe un límite en la forma en que la lámina β puede curvarse para formar el barril (Figura 11-26). Desde este punto de vista, una lámina β es menos versátil que un conjunto de hélices α. Figura Estructura tridimensional de una porina de la membrana externa de una bacteria (Rhodoabacter capsulatus) determinada por cristalografía de rayos X La proteína forma una lámina β de 16 cadenas, que se curva sobre si misma formando un canal transmembrana lleno de agua. Aunque no se muestra en la figura, las porinas se asocian de tres en tres formando trímeros, con tres canales independientes (Adaptado de S.W. Cowan, Curr. Opin Struct. Biol.. 3: , 1993) 36
37 Las proteínas de membrana se pueden solubilizar purificar mediante detergentes Para comprender totalmente una proteína es necesario conocer con todo detalle su estructura, lo cual no es fácil en el caso de las proteínas de membrana. La mayoría de procedimientos bioquímicos han sido diseñados para trabajar con moléculas disueltas en agua o en disolventes sencillos; sin embargo, las proteínas de membrana están diseñadas para trabajar en un ambiente que es parcialmente acuoso y parcialmente lipídico: sacarlas de este ambiente y purificarlas manteniendo su estructura, es un trabajo arduo. Para estudiar detalladamente una proteína determinada es necesario separarla del resto de proteínas. Para la mayoría de proteínas de membrana esta primera etapa del proceso de aislamiento implica la solubilización de la membrana con agentes que destruyen la bicapa lipídica y rompen las asociaciones hidrofóbicas. Para ello los más útiles son los detergentes, moléculas parecidas a lípidos, anfipáticas y pequeñas, con regiones hidrofilicas y regiones hidrofóbicas (véase Figura 11 27). Los detergentes se diferencian de los fosfolípidos de membrana en que disponen de una sola cola hidrofóbica y, por tanto, se comportan de un modo significativamente distinto. Debido a su cola hidrofóbica sencilla, las moléculas de detergente adoptan formas más parecidas a los conos que a los cilindros y en el agua tienden a agregarse en pequeños grupos llamados micelas, en lugar de formar bicapas más cilíndricas, como los fosfolípidos. Al mezclarlos en exceso con las membranas, los extremos hidrofóbicos de las moléculas de detergente se unen a las regiones hidrofóbicas de la zona externa de las proteínas transmembrana, y también a las colas hidrofóbicas de las moléculas de fosfolípido, separándolos de las proteínas. Puesto que el otro extremo de la molécula de detergente es hidrofílico, la unión tiende a disolver las proteínas de membrana en la solución en forma de complejos detergente-proteína (Figura 11 28). Al mismo tiempo, el detergente solubiliza los fosfolípidos. Los complejos proteína detergente pueden separarse unos de otros y de los complejos lípido detergente mediante electroforesis en gel de poliacrilamida con SDS (véase Capítulo 5). 37
38 Figura Estructura de dos detergentes utilizados habitualmente El dodecil sulfato sódico (SDS) es un detergente iónico fuerte (es decir, que tiene un grupo ionizado en su extremo hidrofílico), y el Tritón X- 100, es un detergente no iónico suave (es decir, tiene una estructura no ionizada pero polar en su extremo hidrofílico). La porción hidrofóbica de cada detergente se muestra en verde, y la porción hidrofílica en rojo. Obsérvese que la zona marcada entre corchetes del Tritón X-100 se repite unas ocho veces. Los detergentes iónicos fuertes como el SDS, no sólo desplazan las moléculas lipídicas de las proteínas sino que también producen un desplegamiento de estas proteínas. (Véase Panel 5-5, pp ). 38
39 Figura Utilización de detergentes suaves como el Tritón X 100 para solubilizar proteínas de membrana El detergente rompe la bicapa lipídica e incorpora la proteína a la solución en forma de complejos proteína detergente. Los fosfolípidos de la membrana también se solubilizan por acción del detergente. Como se ilustra en la figura, las moléculas de detergente tienen forma de cono y tienden a agregarse formando grupos llamados micelas 39
40 Se conoce la estructura completa de muy pocas proteínas de membrana Mucho de lo que sabemos acerca de la estructura de las proteínas de membrana lo hemos aprendido a través de medidas indirectas. El método directo estándar para determinar la estructura proteica se basa en la cristalografía de rayos X (se discute en el Capítulo 5), pero requiere que la molécula a estudiar adopte una disposición cristalina ordenada, y es muy difícil lograr que las proteínas transmembrana cristalicen. Sin embargo, hay dos excepciones notables, la bacteriorrodopsina y el centro de reacción fotosintético; ambas son proteínas de membrana bacterianas que desempeñan un papel primordial en la captación y utilización de la energía procedente de la luz solar. La estructura de estas dos proteínas ha revelado exactamente de qué forma las hélices α cruzan la bicapa lipídica, y también cómo pueden ensamblarse distintos tipos de moléculas proteicas formando complejos funcionales en una membrana. La estructura de la bacteriorrodopsina se muestra en la Figura Esta pequeña proteína (de unos 250 aminoácidos) se encuentra en grandes cantidades en la membrana plasmática de la arqueobacteria Halobacterium halobium, en los pantanos salinos. La bacteriorrodopsina actúa como una proteína transportadora que bombea H + hacia el exterior de la bacteria. Este bombeo necesita energía, y la bacteriorrodopsina capta directamente la energía de la luz solar. Cada molécula de bacteriorrodopsina contiene una molécula no proteica llamada retinal que absorbe la luz y que le da a la proteína (y a la bacteria) su característico color púrpura. Esta pequeña molécula hidrofóbica está unida covalentemente a una de las siete hélices α transmembrana de la bacteriorrodopsina, y está situada en el plano de la bicapa lipídica, completamente envuelta por estas siete hélices α (véase Figura 11 29). Cuando el retinal absorbe un fotón de luz, cambia de forma provocando una serie de pequeños cambios conformacionales en la proteína situada en la bicapa lipídica. Estos cambios dan lugar a la transferencia de un H + desde el retinal hasta el exterior de la bacteria: el H + se desplaza a través de la bicapa a lo largo de una vía de cadenas laterales de aminoácidos polares colocados estratégicamente (véase Figura 11 29). Entonces, el retinal se regenera captando un H + del citosol y la proteína recupera su conformación original, con lo cual el ciclo puede reiniciarse. El resultado de todo este proceso es la transferencia de un H + hacia el exterior de la bacteria, lo que disminuye 40
LA MEMBRANA PLASMÁTICA
 ENVOLTURA CELULAR Todas las células tienen que mantener un medio interno adecuado para poder llevar a cabo las reacciones químicas necesarias para la vida. Por ello, están rodeadas de una fina membrana
ENVOLTURA CELULAR Todas las células tienen que mantener un medio interno adecuado para poder llevar a cabo las reacciones químicas necesarias para la vida. Por ello, están rodeadas de una fina membrana
 LA CELULA MEMBRANA PLASMATICA La unidad estructural,histologica y anatomica de los seres vivos es la celula y cada una de ellas se organiza en tejidos, organos y aparatos, orientados a una funcion especifica.
LA CELULA MEMBRANA PLASMATICA La unidad estructural,histologica y anatomica de los seres vivos es la celula y cada una de ellas se organiza en tejidos, organos y aparatos, orientados a una funcion especifica.
Lección 21. Proteínas de membrana Objetivos:
 Lección 21. Proteínas de membrana Objetivos: 1. Enumerar las funciones de las proteínas de membrana 2. Clasificar las proteínas de membrana 3. Describir las interacciones de las proteínas con la bicapa
Lección 21. Proteínas de membrana Objetivos: 1. Enumerar las funciones de las proteínas de membrana 2. Clasificar las proteínas de membrana 3. Describir las interacciones de las proteínas con la bicapa
EL TRANSPORTE CELULAR
 EL TRANSPORTE CELULAR Sumario Historia de la Teoría Celular Estructura y función celular Transporte celular 1. Membrana Celular 2. La Difusión 3. La Osmosis 4. La Difusión Facilitada 5. El Transporte Activo
EL TRANSPORTE CELULAR Sumario Historia de la Teoría Celular Estructura y función celular Transporte celular 1. Membrana Celular 2. La Difusión 3. La Osmosis 4. La Difusión Facilitada 5. El Transporte Activo
Las membranas conforman los límites de las células; están constituidas por una bicapa lipídica
 Células y membranas Las membranas conforman los límites de las células; están constituidas por una bicapa lipídica Los orgánulos: son compartimentos rodeados de membrana situadas en el interior de las
Células y membranas Las membranas conforman los límites de las células; están constituidas por una bicapa lipídica Los orgánulos: son compartimentos rodeados de membrana situadas en el interior de las
El agua como disolvente
 hidrofobicas El agua como disolvente El elevado momento dipolar del agua y su facilidad para formar puentes de hidrógeno hacen que el agua sea un excelente disolvente. Una molécula o ión es soluble en
hidrofobicas El agua como disolvente El elevado momento dipolar del agua y su facilidad para formar puentes de hidrógeno hacen que el agua sea un excelente disolvente. Una molécula o ión es soluble en
MEMBRANAS 2. http://www.changeipadwallpaper.com/wp-content/uploads/1024ipad/scenery/rafts%20landscape.jpg BALSAS Y TRANSPORTE
 MEMBRANAS 2 http://www.changeipadwallpaper.com/wp-content/uploads/1024ipad/scenery/rafts%20landscape.jpg BALSAS Y TRANSPORTE MICRODOMINIOS DE ESFINGOLÍPIDOS Y COLESTEROL MAS GRUESO Y MAS ORDENADO (MENOS
MEMBRANAS 2 http://www.changeipadwallpaper.com/wp-content/uploads/1024ipad/scenery/rafts%20landscape.jpg BALSAS Y TRANSPORTE MICRODOMINIOS DE ESFINGOLÍPIDOS Y COLESTEROL MAS GRUESO Y MAS ORDENADO (MENOS
hidratos de carbono lípidos, proteínas, vitaminas sales minerales y agua principios inmediatos aminoácidos ácidos grasos esenciales
 A pesar de la diversidad en su composición, los análisis químicos revelan que todos los alimentos presentan seis grupos de sustancias comunes, denominadas nutritivas, en distinta proporción: hidratos de
A pesar de la diversidad en su composición, los análisis químicos revelan que todos los alimentos presentan seis grupos de sustancias comunes, denominadas nutritivas, en distinta proporción: hidratos de
Generación y Transmisión de señales en las Neuronas
 Generación y Transmisión de señales en las Neuronas Los detalle de la transmisión sináptica (comunicación entre neuronas) se explicará en el siguiente capítulo. En éste trataremos de los cambios intraneuronal:
Generación y Transmisión de señales en las Neuronas Los detalle de la transmisión sináptica (comunicación entre neuronas) se explicará en el siguiente capítulo. En éste trataremos de los cambios intraneuronal:
Procariota y Eucariota
 Célula Todas las células comparten dos características esenciales. La primera es una membrana externa, la membrana celular -o membrana plasmática- que separa el citoplasma de la célula de su ambiente externo.
Célula Todas las células comparten dos características esenciales. La primera es una membrana externa, la membrana celular -o membrana plasmática- que separa el citoplasma de la célula de su ambiente externo.
Área Académica de: Química. Programa Educativo: Licenciatura de Química en Alimentos. Nombre de la Asignatura: Biología celular
 Área Académica de: Química Línea de Investigación Programa Educativo: Licenciatura de Química en Alimentos Nombre de la Asignatura: Biología celular Tema: Introducción Ciclo: Agosto-Diciembre 2011 Profesor(a):
Área Académica de: Química Línea de Investigación Programa Educativo: Licenciatura de Química en Alimentos Nombre de la Asignatura: Biología celular Tema: Introducción Ciclo: Agosto-Diciembre 2011 Profesor(a):
Evolución de la vida en la tierra:la Célula
 Evolución de la vida en la tierra:la Célula Nuestro planeta tierra no siempre ha sido igual, sin embargo todos los astros que forman el universo están compuestos por los mismos elementos y están controlados
Evolución de la vida en la tierra:la Célula Nuestro planeta tierra no siempre ha sido igual, sin embargo todos los astros que forman el universo están compuestos por los mismos elementos y están controlados
La Absorción del Agua
 La Absorción del Agua Importancia del Agua en las Plantas Es el cons5tuyente principal del protoplasma celular, en ocasiones representa hasta el 95% del peso total de la planta. Es el solvente en el que
La Absorción del Agua Importancia del Agua en las Plantas Es el cons5tuyente principal del protoplasma celular, en ocasiones representa hasta el 95% del peso total de la planta. Es el solvente en el que
Capítulo 5: BIOMOLÉCULAS
 Capítulo 5: BIOMOLÉCULAS De qué están hechas las células? Al analizar los átomos y moléculas presentes en las células, se observa que todas ellas se asemejan: una gran proporción es agua; el resto es un
Capítulo 5: BIOMOLÉCULAS De qué están hechas las células? Al analizar los átomos y moléculas presentes en las células, se observa que todas ellas se asemejan: una gran proporción es agua; el resto es un
Tema 2. El agua, el ph y los equilibrios iónicos
 Tema 2. El agua, el ph y los equilibrios iónicos -Interacciones débiles en sistemas acuosos: puentes de hidrógeno, fuerzas de van der Waals e interacciones iónicas. -Enlaces hidrofóbicos. -Estructura y
Tema 2. El agua, el ph y los equilibrios iónicos -Interacciones débiles en sistemas acuosos: puentes de hidrógeno, fuerzas de van der Waals e interacciones iónicas. -Enlaces hidrofóbicos. -Estructura y
METABOLISMO CELULAR. Es el conjunto de reacciones químicas a través de las cuales el organismo intercambia materia y energía con el medio
 METABOLISMO CELULAR Es el conjunto de reacciones químicas a través de las cuales el organismo intercambia materia y energía con el medio Reacciones Celulares Básicas. Los sistemas vivos convierten la energía
METABOLISMO CELULAR Es el conjunto de reacciones químicas a través de las cuales el organismo intercambia materia y energía con el medio Reacciones Celulares Básicas. Los sistemas vivos convierten la energía
TEMA 8 ESTRUCTURA CRISTALINA
 Tema 8. Estructura cristalina 1 TEMA 8 ESTRUCTURA CRISTALINA Los sólidos pueden clasificarse: 1.- Por su ordenación: 1a. Sólidos amorfos: tienen una estructura desordenada. Sus átomos o moléculas se colocan
Tema 8. Estructura cristalina 1 TEMA 8 ESTRUCTURA CRISTALINA Los sólidos pueden clasificarse: 1.- Por su ordenación: 1a. Sólidos amorfos: tienen una estructura desordenada. Sus átomos o moléculas se colocan
Practica la genética Ficha didáctica del profesorado Bachillerato
 Ficha didáctica del profesorado Bachillerato www.eurekamuseoa.es Extracción de ADN Cuál es la función de cada uno de los ingredientes utilizados para realizar la disolución tampón para la visualización
Ficha didáctica del profesorado Bachillerato www.eurekamuseoa.es Extracción de ADN Cuál es la función de cada uno de los ingredientes utilizados para realizar la disolución tampón para la visualización
Lisofosfolípidos y digestibilidad de grasas
 Lisofosfolípidos y digestibilidad de grasas Pintaluba Fig.1 Micela La grasa se compone principalmente de triglicéridos y no están solubles en agua sin embargo su digestión tiene lugar en un ambiente acuoso.
Lisofosfolípidos y digestibilidad de grasas Pintaluba Fig.1 Micela La grasa se compone principalmente de triglicéridos y no están solubles en agua sin embargo su digestión tiene lugar en un ambiente acuoso.
De manera general la teoría celular moderna se resume en tres postulados:
 De manera general la teoría celular moderna se resume en tres postulados: La célula es la unidad básica estructural de todos los seres vivos, todos los organismos están formados por células. La célula
De manera general la teoría celular moderna se resume en tres postulados: La célula es la unidad básica estructural de todos los seres vivos, todos los organismos están formados por células. La célula
Ablandamiento de agua mediante el uso de resinas de intercambio iónico.
 Ablandamiento de agua por intercambio iónica página 1 Ablandamiento de agua mediante el uso de resinas de intercambio iónico. (Fuentes varias) Algunos conceptos previos: sales, iones y solubilidad. Que
Ablandamiento de agua por intercambio iónica página 1 Ablandamiento de agua mediante el uso de resinas de intercambio iónico. (Fuentes varias) Algunos conceptos previos: sales, iones y solubilidad. Que
Definición de la célula
 Página 1 de 8 La Célula: estructura interna y metabolismo Definición de la célula La célula se entiende como la unidad mínima de un organismo capaz de actuar de manera autónoma en su funcionamiento y reproducción.
Página 1 de 8 La Célula: estructura interna y metabolismo Definición de la célula La célula se entiende como la unidad mínima de un organismo capaz de actuar de manera autónoma en su funcionamiento y reproducción.
I) Biomoléculas 2) El agua I-2 EL AGUA
 Biomoléculas 2) El agua I-2 EL AGUA") I-2 EL AGUA IMPRTANCIA DEL AGUA PARA LS SERES VIVS El agua es el líquido más abundante de la corteza y uno de los pocos líquidos naturales. No es de extrañar entonces que el agua sea una sustancia esencial
I-2 EL AGUA IMPRTANCIA DEL AGUA PARA LS SERES VIVS El agua es el líquido más abundante de la corteza y uno de los pocos líquidos naturales. No es de extrañar entonces que el agua sea una sustancia esencial
ANALOGÍA: Célula vegetal / Fábrica
 ANALOGÍA: Célula vegetal / Fábrica Introducir el tópico. Una célula es un sistema en el que tienen lugar los procesos básicos para realizar sus funciones vitales: nutrición, relación y multiplicación,
ANALOGÍA: Célula vegetal / Fábrica Introducir el tópico. Una célula es un sistema en el que tienen lugar los procesos básicos para realizar sus funciones vitales: nutrición, relación y multiplicación,
CÓDIGO GENÉTICO Y SÍNTESIS DE PROTEÍNAS
 CÓDIGO GENÉTICO Y SÍNTESIS DE PROTEÍNAS Sumario Mitosis y meiosis Código genético y síntesis de proteínas: 1. Concepto de gen 2. Estructura del ADN 3. La replicación del ADN 4. La transcripción 5. La traducción
CÓDIGO GENÉTICO Y SÍNTESIS DE PROTEÍNAS Sumario Mitosis y meiosis Código genético y síntesis de proteínas: 1. Concepto de gen 2. Estructura del ADN 3. La replicación del ADN 4. La transcripción 5. La traducción
Modulo 2 NIVEL CELULAR DE ORGANIZACIÓN DEL CUERPO HUMANO
 Modulo 2 NIVEL CELULAR DE ORGANIZACIÓN DEL CUERPO HUMANO NIVEL CELULAR DE ORGANIZACIÓN DEL CUERPO HUMANO Los seres vivos están integrados por moléculas (inanimadas) los organismos vivos poseen atributos
Modulo 2 NIVEL CELULAR DE ORGANIZACIÓN DEL CUERPO HUMANO NIVEL CELULAR DE ORGANIZACIÓN DEL CUERPO HUMANO Los seres vivos están integrados por moléculas (inanimadas) los organismos vivos poseen atributos
Elementos de Física - Aplicaciones ENERGÍA. Taller Vertical 3 de Matemática y Física Aplicadas MASSUCCO ARRARÁS MARAÑON DI LEO
 Elementos de Física - Aplicaciones ENERGÍA Taller Vertical 3 de Matemática y Física Aplicadas MASSUCCO ARRARÁS MARAÑON DI LEO Energía La energía es una magnitud física que está asociada a la capacidad
Elementos de Física - Aplicaciones ENERGÍA Taller Vertical 3 de Matemática y Física Aplicadas MASSUCCO ARRARÁS MARAÑON DI LEO Energía La energía es una magnitud física que está asociada a la capacidad
TEMA 13. EL METABOLISMO CELULAR 5. PROCESOS DE OXIDO-REDUCCIO E EL METABOLISMO
 TEMA 13. EL METABOLISMO CELULAR 1. METABOLISMO CELULAR: GE ERALIDADES. 2. RUTAS METABÓLICAS. 3. TIPOS DE PROCESOS METABÓLICOS. 4. TIPOS METABOLICOS DE SERES VIVOS. 5. PROCESOS DE OXIDO-REDUCCIO E EL METABOLISMO
TEMA 13. EL METABOLISMO CELULAR 1. METABOLISMO CELULAR: GE ERALIDADES. 2. RUTAS METABÓLICAS. 3. TIPOS DE PROCESOS METABÓLICOS. 4. TIPOS METABOLICOS DE SERES VIVOS. 5. PROCESOS DE OXIDO-REDUCCIO E EL METABOLISMO
ESCUELA SUPERIOR POLITECNICA DEL LITORAL OFICINA DE ADMISIONES CURSO DE NIVELACION DE CARRERA SEGUNDO PARCIAL DE QUIMICA Marzo de 2014
 ESCUELA SUPERIOR POLITECNICA DEL LITORAL OFICINA DE ADMISIONES CURSO DE NIVELACION DE CARRERA SEGUNDO PARCIAL DE QUIMICA Marzo de 2014 NOMBRE: PARALELO: FECHA: El presente examen ha sido elaborado para
ESCUELA SUPERIOR POLITECNICA DEL LITORAL OFICINA DE ADMISIONES CURSO DE NIVELACION DE CARRERA SEGUNDO PARCIAL DE QUIMICA Marzo de 2014 NOMBRE: PARALELO: FECHA: El presente examen ha sido elaborado para
Completar: Un sistema material homogéneo constituido por un solo componente se llama.
 IES Menéndez Tolosa 3º ESO (Física y Química) 1 Completar: Un sistema material homogéneo constituido por un solo componente se llama. Un sistema material homogéneo formado por dos o más componentes se
IES Menéndez Tolosa 3º ESO (Física y Química) 1 Completar: Un sistema material homogéneo constituido por un solo componente se llama. Un sistema material homogéneo formado por dos o más componentes se
Solubilidad. y se representa por.
 Solubilidad Solubilidad. La solubilidad mide la cantidad máxima de soluto capaz de disolverse en una cantidad definida de disolvente, a una temperatura determinada, y formar un sistema estable que se denomina
Solubilidad Solubilidad. La solubilidad mide la cantidad máxima de soluto capaz de disolverse en una cantidad definida de disolvente, a una temperatura determinada, y formar un sistema estable que se denomina
UNIDAD Nº3 BIOLOGÍA CELULAR LA CÉLULA Y SUS COMPONENTES
 UNIDAD Nº3 BIOLOGÍA CELULAR LA CÉLULA Y SUS COMPONENTES OBJETIVOS Comprender el concepto de célula reconociendo sus distintos tipos celulares. Integrar el conocimiento de la estructura de los componentes
UNIDAD Nº3 BIOLOGÍA CELULAR LA CÉLULA Y SUS COMPONENTES OBJETIVOS Comprender el concepto de célula reconociendo sus distintos tipos celulares. Integrar el conocimiento de la estructura de los componentes
Tema 14. La Fase luminosa de la fotosíntesis Javier Corzo. Departamento de Bioquímica y Biología Molecular Universidad de La Laguna
 Tema 14. La Fase luminosa de la fotosíntesis Javier orzo. Departamento de Bioquímica y Biología Molecular Universidad de La Laguna 1. Vamos a considerar como fotosíntesis exclusivamente al proceso de transformación
Tema 14. La Fase luminosa de la fotosíntesis Javier orzo. Departamento de Bioquímica y Biología Molecular Universidad de La Laguna 1. Vamos a considerar como fotosíntesis exclusivamente al proceso de transformación
Preguntas de selectividad en Andalucía. Ácidos nucleicos. Análisis e interpretación de imágenes, esquemas, figuras...
 Año 2001 Describa las funciones más relevantes de los nucleótidos. Cite un ejemplo de nucleótido que participe en cada una de ellas [1,5]. Explique las funciones de los distintos tipos de RNA que participan
Año 2001 Describa las funciones más relevantes de los nucleótidos. Cite un ejemplo de nucleótido que participe en cada una de ellas [1,5]. Explique las funciones de los distintos tipos de RNA que participan
Estructura y propiedades de las proteínas
 Estructura y propiedades de las proteínas Matilde Julián Seguí Introducción Las proteínas son las macromoléculas biológicas más importantes. Hay gran variedad de proteínas y cumplen gran variedad de funciones
Estructura y propiedades de las proteínas Matilde Julián Seguí Introducción Las proteínas son las macromoléculas biológicas más importantes. Hay gran variedad de proteínas y cumplen gran variedad de funciones
Clasificación de los mecanismos de transporte de solutos a través de membranas
 Clasificación de los mecanismos de transporte de solutos a través de membranas 1. Transporte no mediado o difusión simple (Bicapa lipídica) ( G < 0) 2. Transporte mediado (Proteínas de membrana) 2.1. T.
Clasificación de los mecanismos de transporte de solutos a través de membranas 1. Transporte no mediado o difusión simple (Bicapa lipídica) ( G < 0) 2. Transporte mediado (Proteínas de membrana) 2.1. T.
Presentación organizada por José Antonio Pascual Trillo
 Presentación organizada por José Antonio Pascual Trillo La fotosíntesis es un proceso que se desarrolla en dos etapas: Reacciones lumínicas: es un proceso dependiente de la luz (etapa clara), requiere
Presentación organizada por José Antonio Pascual Trillo La fotosíntesis es un proceso que se desarrolla en dos etapas: Reacciones lumínicas: es un proceso dependiente de la luz (etapa clara), requiere
Ruta de las pentosas fosfato
 Ruta de las pentosas fosfato La ruta predominante del catabolismo de la glucosa es la glucólisis para dar piruvato, seguida por la oxidación a CO 2 en el ciclo del ácido cítrico. Un proceso alternativo,
Ruta de las pentosas fosfato La ruta predominante del catabolismo de la glucosa es la glucólisis para dar piruvato, seguida por la oxidación a CO 2 en el ciclo del ácido cítrico. Un proceso alternativo,
Células procariotas y eucariotas
 Células procariotas y eucariotas La teoría celular, establece que todos los seres vivos están constituidos por células y que toda célula proviene de una preexistente. En efecto, desde los minúsculos microorganismos
Células procariotas y eucariotas La teoría celular, establece que todos los seres vivos están constituidos por células y que toda célula proviene de una preexistente. En efecto, desde los minúsculos microorganismos
www.elesapiens.com GLOSARIO DE QUÍMICA ABONO: Es cualquier sustancia orgánica o inorgánica que mejora la calidad de la tierra.
 GLOSARIO DE QUÍMICA ABONO: Es cualquier sustancia orgánica o inorgánica que mejora la calidad de la tierra. ANTIBIÓTICO: Es una sustancia química que mata o impide el crecimiento de ciertas clases de microorganismos
GLOSARIO DE QUÍMICA ABONO: Es cualquier sustancia orgánica o inorgánica que mejora la calidad de la tierra. ANTIBIÓTICO: Es una sustancia química que mata o impide el crecimiento de ciertas clases de microorganismos
TIPOS DE RESTRICCIONES
 RESTRICCIONES: Las restricciones son reglas que determinan la posición relativa de las distintas geometrías existentes en el archivo de trabajo. Para poder aplicarlas con rigor es preciso entender el grado
RESTRICCIONES: Las restricciones son reglas que determinan la posición relativa de las distintas geometrías existentes en el archivo de trabajo. Para poder aplicarlas con rigor es preciso entender el grado
Física de los Procesos Biológicos Curso 2005/6
 Bibliografía: ísica, Kane, Tema 8 ísica de los Procesos Biológicos Curso 2005/6 Grupo 3 TEMA 2 BIOMECÁNICA 2.1 SÓIDO DEORMABE Parte 1 Introducción Vamos a estudiar como los materiales se deforman debido
Bibliografía: ísica, Kane, Tema 8 ísica de los Procesos Biológicos Curso 2005/6 Grupo 3 TEMA 2 BIOMECÁNICA 2.1 SÓIDO DEORMABE Parte 1 Introducción Vamos a estudiar como los materiales se deforman debido
Actividad: Qué es capilaridad?
 Qué es capilaridad? Nivel: 3º medio Subsector: Ciencias físicas Unidad temática: Ver video Capilaridad Actividad: Qué es capilaridad? Los fluidos son un conjunto de moléculas distribuidas al azar que se
Qué es capilaridad? Nivel: 3º medio Subsector: Ciencias físicas Unidad temática: Ver video Capilaridad Actividad: Qué es capilaridad? Los fluidos son un conjunto de moléculas distribuidas al azar que se
El agua. El agua. Se pueden ver muy bien los 4 puentes de hidrógeno que forma la molécula de agua central.
 El agua El agua es una de las sustancias más peculiares conocidas. Es una molécula pequeña y sencilla, pero tras su aparente simplicidad presenta una complejidad sorprendente que la hace especialmente
El agua El agua es una de las sustancias más peculiares conocidas. Es una molécula pequeña y sencilla, pero tras su aparente simplicidad presenta una complejidad sorprendente que la hace especialmente
TABLA DE DECISION. Consideremos la siguiente tabla, expresada en forma genérica, como ejemplo y establezcamos la manera en que debe leerse.
 TABLA DE DECISION La tabla de decisión es una herramienta que sintetiza procesos en los cuales se dan un conjunto de condiciones y un conjunto de acciones a tomar según el valor que toman las condiciones.
TABLA DE DECISION La tabla de decisión es una herramienta que sintetiza procesos en los cuales se dan un conjunto de condiciones y un conjunto de acciones a tomar según el valor que toman las condiciones.
TRATAMIENTO DE CALDERAS SQUIB MEXICO LABORATORIOS DE INVESTIGACIÓN DIVISIÓN TRATAMIENTO DE AGUA
 TRATAMIENTO DE CALDERAS SQUIB MEXICO LABORATORIOS DE INVESTIGACIÓN DIVISIÓN TRATAMIENTO DE AGUA INTRODUCCIÓN Con relación a tratamientos de agua para calderas, se ha estudiado ampliamente en el desarrollo
TRATAMIENTO DE CALDERAS SQUIB MEXICO LABORATORIOS DE INVESTIGACIÓN DIVISIÓN TRATAMIENTO DE AGUA INTRODUCCIÓN Con relación a tratamientos de agua para calderas, se ha estudiado ampliamente en el desarrollo
Capítulo 6. Valoración respiratoria
 498 Capítulo 6. Valoración respiratoria 6.19. La respiración. Intercambio gaseoso y modificaciones durante el esfuerzo 6.19 La respiración. Intercambio gaseoso y modificaciones durante el esfuerzo 499
498 Capítulo 6. Valoración respiratoria 6.19. La respiración. Intercambio gaseoso y modificaciones durante el esfuerzo 6.19 La respiración. Intercambio gaseoso y modificaciones durante el esfuerzo 499
Requisitos del semillero
 Requisitos del semillero La tarea de la cama de siembra es proporcionar a la semilla las condiciones idóneas para una germinación rápida y uniforme. Esto requiere agua, aire, calor y un ambiente libre
Requisitos del semillero La tarea de la cama de siembra es proporcionar a la semilla las condiciones idóneas para una germinación rápida y uniforme. Esto requiere agua, aire, calor y un ambiente libre
Las moléculas de los seres vivos Control de la actividad celular Fuente de energía para las células:
 Las moléculas de los seres vivos Control de la actividad celular Fuente de energía para las células: 1. ATP 2. La respiración celular 3. La fermentación Proceso de fotosíntesis La fuente principal de energía
Las moléculas de los seres vivos Control de la actividad celular Fuente de energía para las células: 1. ATP 2. La respiración celular 3. La fermentación Proceso de fotosíntesis La fuente principal de energía
En el APOENZIMA se distinguen tres tipos de aminoácidos:
 1. Concepto de biocatalizador. Son sustancias que consiguen que las reacciones se realicen a gran velocidad a bajas temperaturas, ya que disminuyen la energía de activación de los reactivos. Pueden ser:
1. Concepto de biocatalizador. Son sustancias que consiguen que las reacciones se realicen a gran velocidad a bajas temperaturas, ya que disminuyen la energía de activación de los reactivos. Pueden ser:
5.1. Organizar los roles
 Marco de intervención con personas en grave situación de exclusión social 5 Organización de la acción 5.1. Organizar los roles Parece que el modelo que vamos perfilando hace emerger un rol central de acompañamiento
Marco de intervención con personas en grave situación de exclusión social 5 Organización de la acción 5.1. Organizar los roles Parece que el modelo que vamos perfilando hace emerger un rol central de acompañamiento
La Célula. Unidad Fundamental delavida
 La Célula Unidad Fundamental delavida La Célula. Unidad Fundamental de la vida El descubrimiento de la célula La teoría celular Estructura de la célula Tipos de células Tipos de células eucariotas Orgánulos
La Célula Unidad Fundamental delavida La Célula. Unidad Fundamental de la vida El descubrimiento de la célula La teoría celular Estructura de la célula Tipos de células Tipos de células eucariotas Orgánulos
CONTROL DE LA ACTIVIDAD CELULAR
 CONTROL DE LA ACTIVIDAD CELULAR Sumario Las Moléculas de los Seres Vivos Control de la actividad celular 1. Las reacciones celulares básicas 2. El control de las reacciones celulares 3. Los modelos de
CONTROL DE LA ACTIVIDAD CELULAR Sumario Las Moléculas de los Seres Vivos Control de la actividad celular 1. Las reacciones celulares básicas 2. El control de las reacciones celulares 3. Los modelos de
DL CH12 Reactor químico combinado
 DL CH12 Reactor químico combinado Introducción La reacción química es la operación unitaria que tiene por objeto distribuir de una forma distinta los átomos de unas moléculas (compuestos reaccionantes
DL CH12 Reactor químico combinado Introducción La reacción química es la operación unitaria que tiene por objeto distribuir de una forma distinta los átomos de unas moléculas (compuestos reaccionantes
Estudio de la evaporación
 Estudio de la evaporación Volumen del líquido Tipo de líquido Superficie del recipiente Altura del recipiente Forma del recipiente Presencia de una sal disuelta Introducción Todos hemos observado que una
Estudio de la evaporación Volumen del líquido Tipo de líquido Superficie del recipiente Altura del recipiente Forma del recipiente Presencia de una sal disuelta Introducción Todos hemos observado que una
Ambientación Universitaria BIOLOGÍA. Guía de Actividades 2014
 1 Ambientación Universitaria BIOLOGÍA Guía de Actividades 2014 2 I.- INTRODUCCIÓN- CONCEPTO DE CIENCIA Y VIDA 1) Lea el siguiente texto. Mencione ejemplos de lo que considere investigación en ciencias
1 Ambientación Universitaria BIOLOGÍA Guía de Actividades 2014 2 I.- INTRODUCCIÓN- CONCEPTO DE CIENCIA Y VIDA 1) Lea el siguiente texto. Mencione ejemplos de lo que considere investigación en ciencias
Bolilla 7: Propiedades de los Líquidos
 Bolilla 7: Propiedades de los Líquidos 1 Bolilla 7: Propiedades de los Líquidos Estudiaremos propiedades de los líquidos, derivadas de las fuerzas de cohesión entre las moléculas que lo componen. Además
Bolilla 7: Propiedades de los Líquidos 1 Bolilla 7: Propiedades de los Líquidos Estudiaremos propiedades de los líquidos, derivadas de las fuerzas de cohesión entre las moléculas que lo componen. Además
UNA NUTRICIÓN SANA Los nutrientes son: hidratos de carbono proteína grasa
 UNA NUTRICIÓN SANA La alimentación se compone de varios nutrientes, vitaminas, minerales y agua. Los nutrientes son: Hidratos de carbono Proteínas Grasas Los hidratos de carbono son la glucosa y los almidones
UNA NUTRICIÓN SANA La alimentación se compone de varios nutrientes, vitaminas, minerales y agua. Los nutrientes son: Hidratos de carbono Proteínas Grasas Los hidratos de carbono son la glucosa y los almidones
P (potencia en watios) = U (tensión eléctrica en voltios) x I (corriente eléctrica en amperios)
 = U (tensión eléctrica en voltios) x I (corriente eléctrica en amperios)") 1) La placa solar Introducción Una célula solar o célula fotovoltaica es un componente electrónico que, expuesto a la luz, genera una energía eléctrica. Las baterías de células están generalmente agrupadas
1) La placa solar Introducción Una célula solar o célula fotovoltaica es un componente electrónico que, expuesto a la luz, genera una energía eléctrica. Las baterías de células están generalmente agrupadas
ALTERACIÓN DE LOS ALIMENTOS
 ALTERACIÓN DE LOS ALIMENTOS Introducción Un alimento está alterado cuando en él se presentan cambios que limitan su aprovechamiento. El alimento alterado tiene modificadas sus características organolépticas
ALTERACIÓN DE LOS ALIMENTOS Introducción Un alimento está alterado cuando en él se presentan cambios que limitan su aprovechamiento. El alimento alterado tiene modificadas sus características organolépticas
PREGUNTAS TEST CORRESPONDIENTES A LOS TEMAS 1 AL 5
 PREGUNTAS TEST CORRESPONDIENTES A LOS TEMAS 1 AL 5 Las preguntas de test que le adjuntamos corresponden a exámenes de las últimas convocatorias. Una vez que finalicen el estudio de los cinco primeros capítulos,
PREGUNTAS TEST CORRESPONDIENTES A LOS TEMAS 1 AL 5 Las preguntas de test que le adjuntamos corresponden a exámenes de las últimas convocatorias. Una vez que finalicen el estudio de los cinco primeros capítulos,
Orbitales híbridos. Cajón de Ciencias
 Orbitales híbridos Cajón de Ciencias Los orbitales híbridos son aquellos que se forman por la fusión de otros orbitales. Estudiarlos es un paso básico para entender la geometría y la estructura de las
Orbitales híbridos Cajón de Ciencias Los orbitales híbridos son aquellos que se forman por la fusión de otros orbitales. Estudiarlos es un paso básico para entender la geometría y la estructura de las
Condensación y ebullición ING Roxsana Romero Ariza Junio 2013
 Condensación y ebullición ING Roxsana Romero Ariza Junio 2013 EBULLICIÓN La transferencia de calor a un líquido en ebullición es muy importante en la evaporación y destilación, así como en otros tipos
Condensación y ebullición ING Roxsana Romero Ariza Junio 2013 EBULLICIÓN La transferencia de calor a un líquido en ebullición es muy importante en la evaporación y destilación, así como en otros tipos
Electrólisis. Electrólisis 12/02/2015
 Electrólisis Dr. Armando Ayala Corona Electrólisis La electrolisis es un proceso mediante el cual se logra la disociación de una sustancia llamada electrolito, en sus iones constituyentes (aniones y cationes),
Electrólisis Dr. Armando Ayala Corona Electrólisis La electrolisis es un proceso mediante el cual se logra la disociación de una sustancia llamada electrolito, en sus iones constituyentes (aniones y cationes),
CARACTERÍSTICAS DEL ESTADO VÍTREO BAJO LA AMPLIA DENOMINACIÓN GENÉRICA DE VIDRIOS O DE CUERPOS VÍTREOS QUEDA COMPRENDIDA UNA GRAN VARIEDAD
 CARACTERÍSTICAS DEL ESTADO VÍTREO BAJO LA AMPLIA DENOMINACIÓN GENÉRICA DE VIDRIOS O DE CUERPOS VÍTREOS QUEDA COMPRENDIDA UNA GRAN VARIEDAD DE SUSTANCIAS QUE, AUNQUE A TEMPERATURA AMBIENTE TIENEN LA APARIENCIA
CARACTERÍSTICAS DEL ESTADO VÍTREO BAJO LA AMPLIA DENOMINACIÓN GENÉRICA DE VIDRIOS O DE CUERPOS VÍTREOS QUEDA COMPRENDIDA UNA GRAN VARIEDAD DE SUSTANCIAS QUE, AUNQUE A TEMPERATURA AMBIENTE TIENEN LA APARIENCIA
Dar a conocer la capacidad de disolución del agua frente a otras sustancias.
 MINISTERIO DE EDUCACION Actividad 1: Agua en la vida II. Laboratorio: Solubilidad del agua 1. Tema: AGUA DISOLVENTE UNIVERSAL 2. Objetivo: Dar a conocer la capacidad de disolución del agua frente a otras
MINISTERIO DE EDUCACION Actividad 1: Agua en la vida II. Laboratorio: Solubilidad del agua 1. Tema: AGUA DISOLVENTE UNIVERSAL 2. Objetivo: Dar a conocer la capacidad de disolución del agua frente a otras
ES 1 097 480 U ESPAÑA 11. Número de publicación: 1 097 480. Número de solicitud: 201331388 A47G 29/00 (2006.01) 03.12.2013
 03.12.2013") 19 OFICINA ESPAÑOLA DE PATENTES Y MARCAS ESPAÑA 11 21 Número de publicación: 1 097 480 Número de solicitud: 1331388 1 Int. CI.: A47G 29/00 (06.01) 12 SOLICITUD DE MODELO DE UTILIDAD U 22 Fecha de presentación:
19 OFICINA ESPAÑOLA DE PATENTES Y MARCAS ESPAÑA 11 21 Número de publicación: 1 097 480 Número de solicitud: 1331388 1 Int. CI.: A47G 29/00 (06.01) 12 SOLICITUD DE MODELO DE UTILIDAD U 22 Fecha de presentación:
Modelo corpuscular elemental de la materia
 Modelo corpuscular elemental de la materia Todo lo que es materia sean sólidos, líquidos o gases, está constituida por átomos y moléculas. Sin embargo el diferente aspecto que observamos de una misma sustancia,
Modelo corpuscular elemental de la materia Todo lo que es materia sean sólidos, líquidos o gases, está constituida por átomos y moléculas. Sin embargo el diferente aspecto que observamos de una misma sustancia,
III. DIFUSION EN SOLIDOS
 Metalografía y Tratamientos Térmicos III - 1 - III. DIFUSION EN SOLIDOS III.1. Velocidad de procesos en sólidos Muchos procesos de producción y aplicaciones en materiales de ingeniería están relacionados
Metalografía y Tratamientos Térmicos III - 1 - III. DIFUSION EN SOLIDOS III.1. Velocidad de procesos en sólidos Muchos procesos de producción y aplicaciones en materiales de ingeniería están relacionados
FISIOLOGÍA GENERAL Jesús Merino Pérez y María José Noriega Borge
 EL AGUA: VOLÚMENES Y COMPOSICIÓN DE LOS LÍQUIDOS CORPORALES La vida comenzó en el agua del mar, y las condiciones que reinaban en aquel ambiente primigenio marcaron las características químicas de la materia
EL AGUA: VOLÚMENES Y COMPOSICIÓN DE LOS LÍQUIDOS CORPORALES La vida comenzó en el agua del mar, y las condiciones que reinaban en aquel ambiente primigenio marcaron las características químicas de la materia
MICROECONOMÍA II. PRÁCTICA TEMA II: Equilibrio parcial
 MICROECONOMÍA II PRÁCTICA TEMA II: Equilibrio parcial EJERCICIO 1 A) En equilibrio, la cantidad demandada coincide con la cantidad ofrecida, así como el precio de oferta y demanda. Por lo tanto, para hallar
MICROECONOMÍA II PRÁCTICA TEMA II: Equilibrio parcial EJERCICIO 1 A) En equilibrio, la cantidad demandada coincide con la cantidad ofrecida, así como el precio de oferta y demanda. Por lo tanto, para hallar
TRABAJO PRÁCTICO: MODELOS EXTREMOS DE SUSTANCIA
 1 Complemento TRABAJO PRÁCTICO: S EXTREMOS DE SUSTANCIA FUNDAMENTOS TEÓRICOS BÁSICOS Todas las sustancias pueden considerarse formadas por un conjunto de unidades estructurales o fundamentales. La clase
1 Complemento TRABAJO PRÁCTICO: S EXTREMOS DE SUSTANCIA FUNDAMENTOS TEÓRICOS BÁSICOS Todas las sustancias pueden considerarse formadas por un conjunto de unidades estructurales o fundamentales. La clase
CONCEPTOS DE LA FUERZA
 CONCEPTOS DE LA FUERZA PAPEL DE LA FUERZA EN EL RENDIMIENTO DEPORTIVO La mejora de la fuerza es un factor importante en todas las actividades deportivas, y en algunos casos determinantes (en el arbitraje
CONCEPTOS DE LA FUERZA PAPEL DE LA FUERZA EN EL RENDIMIENTO DEPORTIVO La mejora de la fuerza es un factor importante en todas las actividades deportivas, y en algunos casos determinantes (en el arbitraje
LABORATORIO DE QUÍMICA FACULTAD DE FARMACIA CRISTALIZACIÓN.
 CRISTALIZACIÓN. Un compuesto orgánico cristalino está constituido por un empaquetamiento tridimensional de moléculas unidas principalmente por fuerzas de Van der Waals, que originan atracciones intermoleculares
CRISTALIZACIÓN. Un compuesto orgánico cristalino está constituido por un empaquetamiento tridimensional de moléculas unidas principalmente por fuerzas de Van der Waals, que originan atracciones intermoleculares
35 Facultad de Ciencias Universidad de Los Andes Mérida-Venezuela. Potencial Eléctrico
 q 1 q 2 Prof. Félix Aguirre 35 Energía Electrostática Potencial Eléctrico La interacción electrostática es representada muy bien a través de la ley de Coulomb, esto es: mediante fuerzas. Existen, sin embargo,
q 1 q 2 Prof. Félix Aguirre 35 Energía Electrostática Potencial Eléctrico La interacción electrostática es representada muy bien a través de la ley de Coulomb, esto es: mediante fuerzas. Existen, sin embargo,
Partes de los Espermatozoide
 PRÁCTICA DE LABORATORIO 5 OBSERVACIÓN DE CÉLULAS SEXUALES MASCULINAS Docente: Rolando Hernández Lazo INTRODUCCIÓN Espermatozoide es la célula reproductora sexual masculina o gameto masculino encargada
PRÁCTICA DE LABORATORIO 5 OBSERVACIÓN DE CÉLULAS SEXUALES MASCULINAS Docente: Rolando Hernández Lazo INTRODUCCIÓN Espermatozoide es la célula reproductora sexual masculina o gameto masculino encargada
Illustrator CS4: guía de maquetación
 Paso 1 de 12 En este tutorial vamos a ver cómo hacer un banner de página web. Al hacer este banner de página web aprenderemos varias técnicas de dibujo, como la utilización de la herramienta Elipse, la
Paso 1 de 12 En este tutorial vamos a ver cómo hacer un banner de página web. Al hacer este banner de página web aprenderemos varias técnicas de dibujo, como la utilización de la herramienta Elipse, la
A continuación se presenta los resultados obtenidos en las pruebas realizadas en
 6.0 RESULTADOS, COMPARACIÓN Y ANALISIS. 6.1 PERMEABILIDAD. A continuación se presenta los resultados obtenidos en las pruebas realizadas en el laboratorio para la determinación del coeficiente de permeabilidad
6.0 RESULTADOS, COMPARACIÓN Y ANALISIS. 6.1 PERMEABILIDAD. A continuación se presenta los resultados obtenidos en las pruebas realizadas en el laboratorio para la determinación del coeficiente de permeabilidad
2º E.S.O. - CIENCIAS DE LA NATURALEZA
 2º E.S.O. - CIENCIAS DE LA NATURALEZA En la actualidad se conocen más de 1.800.000 especies de seres vivos distintos (hay muchas otras aun por descubrir). Los seres vivos se diferencian de la materia inerte
2º E.S.O. - CIENCIAS DE LA NATURALEZA En la actualidad se conocen más de 1.800.000 especies de seres vivos distintos (hay muchas otras aun por descubrir). Los seres vivos se diferencian de la materia inerte
Envolturas celulares
 Envolturas celulares LA MEMBRANA PLASMÁTICA Es una envoltura continua, fina y flexible, de unos 6 a 10 nm de grosor, que rodea a la célula, separándo su interior del medio externo, permitiéndole mantener
Envolturas celulares LA MEMBRANA PLASMÁTICA Es una envoltura continua, fina y flexible, de unos 6 a 10 nm de grosor, que rodea a la célula, separándo su interior del medio externo, permitiéndole mantener
EQUILIBRIO QUÍMICO: REACCIONES ÁCIDO-BASE
 Página: 1/7 DEPARTAMENTO ESTRELLA CAMPOS PRÁCTICO 8: EQUILIBRIO QUÍMICO: REACCIONES ÁCIDO-BASE Bibliografía: Química, La Ciencia Central, T.L. Brown, H. E. LeMay, Jr., B. Bursten; Ed. Prentice-Hall, Hispanoamérica,
Página: 1/7 DEPARTAMENTO ESTRELLA CAMPOS PRÁCTICO 8: EQUILIBRIO QUÍMICO: REACCIONES ÁCIDO-BASE Bibliografía: Química, La Ciencia Central, T.L. Brown, H. E. LeMay, Jr., B. Bursten; Ed. Prentice-Hall, Hispanoamérica,
La energía y sus transformaciones
 La energía y sus transformaciones Índice 1 Definición de energía 2 Energías renovables y no renovables 2.1 Energías no renovables 2.2 Energías renovables 3 Transformaciones energéticas 4 Conservación de
La energía y sus transformaciones Índice 1 Definición de energía 2 Energías renovables y no renovables 2.1 Energías no renovables 2.2 Energías renovables 3 Transformaciones energéticas 4 Conservación de
FUNDAMENTOS BIOLÓGICOS DEL APRENDIZAJE Y LA MEMORIA
 Departamento de Biología a Ambiental y Salud PúblicaP FUNDAMENTOS BIOLÓGICOS DEL APRENDIZAJE Y LA MEMORIA La base celular y físico-química del impulso y transmisión de la señal nerviosa. La neurona. Potencial
Departamento de Biología a Ambiental y Salud PúblicaP FUNDAMENTOS BIOLÓGICOS DEL APRENDIZAJE Y LA MEMORIA La base celular y físico-química del impulso y transmisión de la señal nerviosa. La neurona. Potencial
Transporte a través de membranas
 Transporte a través de membranas Javier Corzo. Departamento de Bioquímica y Biología Molecular. Universidad de La Laguna 1 DEFINICIÓN DE TRANSPORTE Consideraremos transporte al movimiento de moléculas
Transporte a través de membranas Javier Corzo. Departamento de Bioquímica y Biología Molecular. Universidad de La Laguna 1 DEFINICIÓN DE TRANSPORTE Consideraremos transporte al movimiento de moléculas
CAPÍTULO 4 INTERACCIÓN DE LA RADIACIÓN CON EL TEJIDO BIOLÓGICO
 CAPÍTULO 4 INTERACCIÓN DE LA RADIACIÓN CON EL TEJIDO BIOLÓGICO El tejido biológico está formado por células, la radiación al interaccionar con el tejido puede tener como blanco de interacción cualquiera
CAPÍTULO 4 INTERACCIÓN DE LA RADIACIÓN CON EL TEJIDO BIOLÓGICO El tejido biológico está formado por células, la radiación al interaccionar con el tejido puede tener como blanco de interacción cualquiera
TEMARIO PARA EL EXAMEN DE BIOQUÍMICA
 TEMARIO PARA EL EXAMEN DE BIOQUÍMICA PARTE I. ESTRUCTURA BIOLÓGICA Y QUÍMICA DE LAS PROTEÍNAS 1. CARACTERÍSTICAS QUÍMICAS DE LA MATERIA VIVA 1. La mayoría de los organismos están compuestos solamente de
TEMARIO PARA EL EXAMEN DE BIOQUÍMICA PARTE I. ESTRUCTURA BIOLÓGICA Y QUÍMICA DE LAS PROTEÍNAS 1. CARACTERÍSTICAS QUÍMICAS DE LA MATERIA VIVA 1. La mayoría de los organismos están compuestos solamente de
Funciones de la membrana plasmática. Tipos de células. Células emisoras
 Funciones de la membrana plasmática. Podemos señalar dos funciones principales: Intercambio de sustancias. La membrana va a dejar pasar hacia el citoplasma determinados nutrientes. Para poder llevar a
Funciones de la membrana plasmática. Podemos señalar dos funciones principales: Intercambio de sustancias. La membrana va a dejar pasar hacia el citoplasma determinados nutrientes. Para poder llevar a
EL FONDO DE MANIOBRA Y LAS NECESIDADES OPERATIVAS DE FONDOS
 2 EL FONDO DE MANIOBRA Y LAS NECESIDADES OPERATIVAS DE FONDOS Las inversiones de una empresa están reflejadas en su activo. Una forma de clasificación de las diferentes inversiones es en función del plazo
2 EL FONDO DE MANIOBRA Y LAS NECESIDADES OPERATIVAS DE FONDOS Las inversiones de una empresa están reflejadas en su activo. Una forma de clasificación de las diferentes inversiones es en función del plazo
Instalaciones de tratamiento de agua de alimentación de caldera
 Instalaciones de tratamiento de agua de alimentación de caldera Introducción La calidad del agua de alimentación a la caldera repercute directamente sobre el buen funcionamiento de la misma así como sobre
Instalaciones de tratamiento de agua de alimentación de caldera Introducción La calidad del agua de alimentación a la caldera repercute directamente sobre el buen funcionamiento de la misma así como sobre
Práctica 1A Ensayo de Granulometría Prácticas de Laboratorio
 1A ENSAYO DE GRANULOMETRÍA 1. TIPOS DE SUELO. RECONOCIMIENTO VISUAL Desde un punto de vista geotécnico, existen cuatro grandes tipos de suelos: gravas, arenas, limos y arcillas, caracterizados principalmente
1A ENSAYO DE GRANULOMETRÍA 1. TIPOS DE SUELO. RECONOCIMIENTO VISUAL Desde un punto de vista geotécnico, existen cuatro grandes tipos de suelos: gravas, arenas, limos y arcillas, caracterizados principalmente
ÁCIDOS NUCLEICOS. Por: Wilfredo Santiago
 ÁCIDOS NUCLEICOS Por: Wilfredo Santiago Ácidos Nucleicos Formados por subunidades llamadas nucleótidos; pueden ser un solo nucleótido o una cadena larga de nucleótidos. Ácidos Nucleicos Nucleótidos individuales:
ÁCIDOS NUCLEICOS Por: Wilfredo Santiago Ácidos Nucleicos Formados por subunidades llamadas nucleótidos; pueden ser un solo nucleótido o una cadena larga de nucleótidos. Ácidos Nucleicos Nucleótidos individuales:
TEMA 5: MODELOS DE ORGANIZACIÓN CELULAR: PROCARIÓTICA Y EUCARIÓTICA (VEGETAL Y ANIMAL)
") TEMA 5: MODELOS DE ORGANIZACIÓN CELULAR: PROCARIÓTICA Y EUCARIÓTICA (VEGETAL Y ) 1.- Describir y diferenciar los dos tipos de organización celular. Comparar las características de las células vegetales
TEMA 5: MODELOS DE ORGANIZACIÓN CELULAR: PROCARIÓTICA Y EUCARIÓTICA (VEGETAL Y ) 1.- Describir y diferenciar los dos tipos de organización celular. Comparar las características de las células vegetales
SISTEMAS DE PRODUCCIÓN
 SISTEMAS DE PRODUCCIÓN La producción es el proceso mediante el cual la empresa transforma un conjunto de factores de producción en un producto cuyo valor debe ser mayor que la suma de los valores de los
SISTEMAS DE PRODUCCIÓN La producción es el proceso mediante el cual la empresa transforma un conjunto de factores de producción en un producto cuyo valor debe ser mayor que la suma de los valores de los
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA
 UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA PROGRAMA DE INGENIERIA DE ALIMENTOS 211612 TRANSFERENCIA DE MASA ACTIVIDAD 11 RECONOCIMIENTO UNIDAD 3 BOGOTA D.C. Extracción líquido - líquido La extracción líquido-líquido,
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA PROGRAMA DE INGENIERIA DE ALIMENTOS 211612 TRANSFERENCIA DE MASA ACTIVIDAD 11 RECONOCIMIENTO UNIDAD 3 BOGOTA D.C. Extracción líquido - líquido La extracción líquido-líquido,
04/11/2013 ALCOHOLES Y FENOLES
 ALCOHOLES Y FENOLES Pueden ser considerados como derivados del agua. Tienen en común, la presencia del grupo funcional OH, conocido como grupo hidroxilo ú oxhídrilo. ALCOHOLES: grupo OH-unido a radical
ALCOHOLES Y FENOLES Pueden ser considerados como derivados del agua. Tienen en común, la presencia del grupo funcional OH, conocido como grupo hidroxilo ú oxhídrilo. ALCOHOLES: grupo OH-unido a radical
COMPOSICIÓN QUÍMICA DE LA CÉLULA.
 COMPOSICIÓN QUÍMICA DE LA CÉLULA. El 99% del peso de una célula está dominado por 6 elementos químicos: carbono, hidrógeno, nitrógeno, oxígeno, fósforo y azufre. La química de los seres vivos, objeto de
COMPOSICIÓN QUÍMICA DE LA CÉLULA. El 99% del peso de una célula está dominado por 6 elementos químicos: carbono, hidrógeno, nitrógeno, oxígeno, fósforo y azufre. La química de los seres vivos, objeto de
Cap. 24 La Ley de Gauss
 Cap. 24 La Ley de Gauss Una misma ley física enunciada desde diferentes puntos de vista Coulomb Gauss Son equivalentes Pero ambas tienen situaciones para las cuales son superiores que la otra Aquí hay
Cap. 24 La Ley de Gauss Una misma ley física enunciada desde diferentes puntos de vista Coulomb Gauss Son equivalentes Pero ambas tienen situaciones para las cuales son superiores que la otra Aquí hay
Láminas epiteliales y uniones célula a célula
 Láminas epiteliales y uniones célula a célula Tomado y modificado de ALBERTS. B. BRAY D. JOHNSON A. LEWIS J. RAFF M. ROBERTS K. WALTER P.: Introducción a la Biología Celular (2ª edición 2006) Editorial
Láminas epiteliales y uniones célula a célula Tomado y modificado de ALBERTS. B. BRAY D. JOHNSON A. LEWIS J. RAFF M. ROBERTS K. WALTER P.: Introducción a la Biología Celular (2ª edición 2006) Editorial
Unidad II Sistemas Dispersos Elaborado por: Q.F.B. Guadalupe Echeagaray Herrera
 Química II (Química General y Orgánica) Unidad II Sistemas Dispersos Elaborado por: Sistemas Dispersos istemas Dispersos: Están constituidos por dos o más sustancias puras, unidas físicamente, (mezcladas).
Química II (Química General y Orgánica) Unidad II Sistemas Dispersos Elaborado por: Sistemas Dispersos istemas Dispersos: Están constituidos por dos o más sustancias puras, unidas físicamente, (mezcladas).