TESIS DOCTORAL: "GANANCIAS Y PÉRDIDAS GENÓMICAS EN TUMORES DE CÁNCER DE MAMA CON FENOTIPO TRIPLE NEGATIVO Y PÉRDIDA DE EXPRESIÓN DE BRCA1.
|
|
|
- Pedro Gutiérrez Quiroga
- hace 6 años
- Vistas:
Transcripción

1 Pontificia Universidad Católica de Chile Facultad de Ciencias Biológicas Programa de Doctorado en Ciencias Biológicas Mención Genética Molecular y Microbiología TESIS DOCTORAL: "GANANCIAS Y PÉRDIDAS GENÓMICAS EN TUMORES DE CÁNCER DE MAMA CON FENOTIPO TRIPLE NEGATIVO Y PÉRDIDA DE EXPRESIÓN DE BRCA1 Por TERESA MARLOREN TAPIA ESPINOZA Agosto 2015
2 Pontificia Universidad Católica de Chile Facultad de Ciencias Biológicas Programa de Doctorado en Ciencias Biológicas Mención Genética Molecular y Microbiología "GANANCIAS Y PÉRDIDAS GENÓMICAS EN TUMORES DE CÁNCER DE MAMA CON FENOTIPO TRIPLE NEGATIVO Y PÉRDIDA DE EXPRESIÓN DE BRCA1 Tesis presentada a la Pontificia Universidad Católica de Chile como parte de los requisitos para optar al grado de Doctor en Ciencias Biológicas mención Genética Molecular y Microbiología Por TERESA MARLOREN TAPIA ESPINOZA Director de Tesis: Comisión de Tesis: Dra. Pilar Carvallo de S-Q. Dr. Francisco Melo Dr. Gareth Owen Dra. Claudia Sáez Agosto 2015
3
4 AGRADECIMIENTOS Comenzaré, dándole las gracias a mí tutora, Dra. Pilar Carvallo, por su formación y apoyo en el desarrollo y discusión de esta memoria de investigación. Además, por su comprensión, tolerancia y espera de mis procesos internos y personales, motivándome para terminar la escritura de este trabajo. Agradezco infinitamente a aquellas mujeres que accedieron a participar de este estudio, en beneficio de la ciencia en Chile y del conocimiento del cáncer de mama. Gracias a mis compañeros y amigos del laboratorio por sus criticas y apoyo, tanto en lo profesional como personal. Especialmente, agradezco a Carolina Álvarez con quien he aclarado muchas dudas respecto a esta memoria de investigación. Gracias a todos por los momentos de entretención durante mi estancia en el laboratorio. Agradezco a quienes financiaron esta tesis: a VRAID-UC, CONICYT, y la Facultad de Ciencias Biológicas de la Pontificia Universidad Católica de Chile, por las becas que me otorgaron para continuar con mi formación científica. A FONDECYT y la beca de Apoyo para la realización de la tesis doctoral de CONICYT por el financiamiento de este proyecto. Asimismo, quiero agradecer a UICC-ICRETT y MECESUP por financiar mi estadía de investigación en el NIH. Y finalmente, a la beca AVON y AACR por permitirme la asistencia a congresos internacionales. Finalmente, gracias a mi familia, mi mamá y papá por todo su apoyo y amor incondicional hacia mí y su nieto. A mis hermanos, por su apoyo y amor a su sobrino. A mí querido hijo Oscar por mostrarme que la capacidad de asombroso aún existe en mí. Gracias a Dios y la virgen María por estar junto a mí y mi familia.
5 A mi madre, padre y Mi querido hijo Oscar
6 INDICE INDICE DE MATERIAS i INDICE DE FIGURAS vi INDICE DE TABLAS viii ABREVIATURAS ix RESUMEN xiii ABSTRACT xvi INTRODUCCION El cáncer de mama Genes de alto riesgo al cáncer de mama hereditario (BRCA1 y BRCA2) Clasificación del cáncer de mama, tanto de esporádico y hereditario, en relación a la histología e inmunofenotipo Definición de los tumores de mama con fenotipo triple negativo Clasificación molecular de los tumores de mama, basada en los perfiles de expresión génica Estructura de la glándula mamaria, y las células epiteliales que la forman Identificación de los tumores de mama tipo Basal mediante inmunohistoquímica El grupo de tumores de mama triple negativo integra tumores del subgrupo tipo Basal Tumores de pacientes portadoras de mutación en el gen BRCA1 se relacionan con el fenotipo triple negativo y el tipo Basal Pérdida de expresión y mecanismos de pérdida de función de BRCA Localización de la proteína BRCA1 en los tumores de mama i
7 12. Funciones de la proteína BRCA1 y su relación con la inestabilidad genómica en los tumores de mama Perfiles de rearreglos genómicos, mediante array-cgh, en cáncer de mama Rearreglos genómicos en tumores de mama triple negativo y tumores del grupo tipo Basal Hipótesis Objetivo General...28 Objetivos específicos PACIENTES Pacientes del estudio Selección de las pacientes con tumores de mama triple negativo Criterios para seleccionar familias con cáncer de mama hereditario MATERIALES Material químico, enzimas y anticuerpos METODOS Análisis de la expresión de las citoqueratinas 5, 14, y del receptor 1 de EGF y de la proteína BRCA1 en tumores TNBC mediante inmunohistoquímica Interpretación de la detección de las citoqueratinas, de EGFR y de la proteína BRCA1 en los tumores TNBC..38 a. Tumores positivos para las CK5 y CK b. Tumores positivos para EGFR c. Tumores positivos para BRCA Valorización de la tinción, calculando el puntaje-h Definición de los tumores TNBC, en tipo basal y no tipo basal ii
8 3.2. Preparación del DNA genómico desde células tumorales Identificación de las células tumorales y microdisección a. Microdisección manual de las células tumorales b. Microdisección por captura con láser de células tumorales Extracción del DNA genómico desde células tumorales microdisectadas manualmente Extracción de DNA genómico desde células tumorales microdisectadas por captura con láser Hibridación genómica comparativa en microarreglos de DNA (array-cgh) Amplificación del DNA genómico tumoral Digestión enzimática del DNA genómico Marcación del DNA genómico digerido Hibridación de las muestras Análisis e identificación de los cambios en el número de copias en el genoma RESULTADOS Definición de los tumores TNBC en tipo Basal y tipo no-basal, por Inmunohistoquímica Análisis de la expresión y localización de la proteína BRCA1 en la glándula mamaria normal y los tumores TNBC Análisis de la expresión de la proteína BRCA1 en los tumores con mutación en BRCA1 o BRCA2 en la línea germinal Expresión y localización de BRCA1, según la clasificación tipo Basal y tipo no-basal de los tumores TNBC iii
9 4.3. Hibridación genómica comparativa en microarreglos de DNA de los tumores de cáncer de mama triple negativo Porcentaje de sondas con alteración de cada cromosoma, detectado en el microarreglo de oligonucleótido de DNA, entre los tumores TNBC Control interno de ganancia, cromosoma X, en los tumores TNBC Análisis de las sondas que reconocen el gen BRCA1, en el microarreglo de oligonucleótido de DNA Análisis del perfil de frecuencia de los cambios en el número de copias del DNA en los tumores TNBC Identificación de los rearreglos genómicos de ganancia y de deleción recurrentes en los tumores TNBC Cambios en el número de copias del DNA diferencia los tumores TNBC/tipo Basal de los TNBC/tipo no-basal Agrupamiento jerárquico no supervisado en base a los genes con ganancia y deleción identificados en los tumores TNBC Análisis de los perfiles de rearreglos genómicos, en relación a la expresión y localización de BRCA1 en los tumores de cáncer de mama triple negativo Regiones genómicas de ganancia y de deleción recurrentes en los tumores agrupados por la expresión y localización de BRCA DISCUSION Los tumores TNBC principalmente se clasifican en el subgrupo tipo Basal, mediante inmunohistoquímica Diferentes patrones de expresión de los marcadores tipo Basal fueron observados entre los tumores TNBC/tipo Basal iv
10 5.3. Diferentes patrones de expresión y localización de la proteína BRCA1 se identifican en los tumores TNBC Perfil de rearreglos genómicos de ganancia y deleciones de los tumores de cáncer de mama triple negativo Perfil de los rearreglos genómicos de tumores de cáncer de mama triple negativo en relación a la expresión y localización de la proteína BRCA CONCLUSIONES REFERENCIA v
11 INDICE DE FIGURAS FIGURA 1: Agrupamiento jerárquico de los tumores de cáncer de mama en 6 subtipos diferentes....8 FIGURA 2: Curvas de Kaplan-Meier de la sobrevida total y de la sobrevida libre de recurrencia de 49 pacientes con cáncer de mama, basada en la clasificación de los diferentes perfiles de expresión génica....9 FIGURA 3: Representación esquemática de la glándula mamaria normal, las estructuras y los tipos de células epiteliales que la forman...12 FIGURA 4: Representación esquemática de la técnica de Hibridación Genómica Comparativa en microarreglos de DNA o array-cgh FIGURA 5: Expresión de las citoqueratinas 5, 14, y EGFR en los tumores TNBC...58 FIGURA 6: Imágenes de la inmunohistoquímica de BRCA1 en tejidos de mama normal, provenientes de reducciones mamarias..72 FIGURA 7: Imágenes de la expresión y localización de BRCA1 en los tumores TNBC FIGURA 8: Gráfico de los tres grupos principales de expresión y localización de la proteína BRCA FIGURA 9: Gráfico de la expresión de BRCA1 en los tumores de cáncer de mama triple negativo 84 FIGURA 10: Porcentaje de sondas con cambio en el número de copias en cada cromosoma y la densidad de genes descrita para cada uno de ellos FIGURA 11: Visualización del cromosoma X en los tumores TNBC vi
12 FIGURA 12: Gráfico de deleción de las 4 sondas dirigidas al gen BRCA1, en la plataforma de microarreglo de oligonucleótido de DNA FIGURA 13: Perfil de frecuencia de los cambios en el número de copias del DNA..97 FIGURA 14: Número de intervalos genómicos identificados en cada tumor TNBC FIGURA 15: Regiones genómicas en estado de ganancia en los tumores TNBC FIGURA 16: Regiones genómicas en estado de deleción en los tumores TNBC FIGURA 17: A) Agrupamiento jerárquico no supervisado de los tumores TNBC, en base la ganancia y deleción de genes alterados en su número de copias. B) Gráfico de la sobrevida total de los casos con Bajo CNA y Alto CNA FIGURA 18: Agrupamiento jerárquico no supervisado de los tumores TNBC, en relación a la expresión y localización de BRCA FIGURA 19: Perfil de frecuencia de los cambios en el número de copias en relación a la expresión y localización de BRCA1 116 vii
13 INDICE DE TABLAS Tabla 1: Características clínico-patológicas de las pacientes chilenas y de sus tumores TNBC, ingresadas al estudio Tabla 2: Características clínicas e histológicas de las pacientes y sus respectivos tumores de cáncer de mama triple negativo. 54 Tabla 3: Expresión de los marcadores tipo Basal citoqueratinas 5, 14 y de EGFR en los tumores de cáncer de mama triple negativo Tabla 4: Tumores de cáncer de mama triple negativo con ganancia de la región cromosómica 17q21.31 que contiene el gen BRCA Tabla 5: Tumores de cáncer de mama triple negativo con deleción de al menos 2 sondas consecutivas del gen BRCA1, en relación a la expresión y localización de la proteína BRCA viii
14 ABREVIATURAS AACS : acetoacetyl-coa synthetase ABL1 : ABL proto-oncogene 1, non-receptor tyrosine kinase ACVR1B : activin A receptor, type IB ADM-1 : Aberration Detection Method-1 AEC : cromógeno 3-amino-9-etilcarbazol AKT1 : v-akt murine thymoma viral oncogene homolog 1 ANAPC5 : anaphase promoting complex subunit 5 ANGPTL7 : angiopoietin-like 7 ATM : ataxia telangiectasia mutated ATXN2 : ataxin 2 BARD1 : BRCA1 associated RING domain 1 BAZ1B : bromodomain adjacent to zinc finger domain, 1B BCL11B : B-cell CLL/lymphoma 11B (zinc finger protein) BRCA1 : Breast cancer susceptibility gene 1 BRCA2 : Breast cancer susceptibility gene 2 BRIP1 : Brca1 Interacting Protein 1 BUB3 : BUB3 mitotic checkpoint protein C12orf43 : chromosome 12 open reading frame 43 CARD16 : caspase recruitment domain family, member 16 CARD18 : caspase recruitment domain family, member 18 CASP1 : caspase 1, apoptosis-related cysteine peptidase CASP4 : caspase 4, apoptosis-related cysteine peptidase CASP5 : caspase 5, apoptosis-related cysteine peptidase CBL : Cbl proto-oncogene, E3 ubiquitin protein ligase CCND1 : cyclin D1 CDCA8 : cell division cycle associated 8 CDK2AP1 : cyclin-dependent kinase 2 associated protein 1 CGH : Hibridación genómica comparative CHEK2 : Cell Cycle Checkpoint Kinase 2 CHFR : checkpoint with forkhead and ring finger domains, E3 ubiquitin protein ligase CIT : citron rho-interacting serine/threonine kinase CK14 : citoqueratinas 14 CK17 : citoqueratinas 17 CK18 : citoqueratinas 18 CK19 : citoqueratinas 19 CK5 : citoqueratinas 5 CK7 : citoqueratinas 7 CK8 : citoqueratinas 8 CLIP1 : CAP-GLY domain containing linker protein 1 CSF1R : colony stimulating factor 1 receptor ix
15 Cy3 : Cyanine 3 Cy5 : Cyanine 5 DAB : cromógeno 3,3-Diaminobenzidina DNA : Acido desoxirribonucleico DSB : Double Strand Breaks EEFSEC : eukaryotic elongation factor, selenocysteine-trna-specific EGFR : Receptor 1 del factor de crecimiento epidermal EIF2B1 : eukaryotic translation initiation factor 2B, subunit 1 alpha, 26kDa EIF4B : eukaryotic translation initiation factor 4B ERBB2 : erb-b2 receptor tyrosine kinase 2 FAM107B : family with sequence similarity 107, member B FDR : False Discovery Rate FFPE : tejido fijado en formalina y embebido en parafina FGFR1 : fibroblast growth factor receptor 1 FGFR2 : fibroblast growth factor receptor 2 FGFR3 : fibroblast growth factor receptor 3 FLT4 : fms-related tyrosine kinase 4 FRAP1 : mechanistic target of rapamycin(serine/threonine kinase) GATA2 : GATA binding protein 2 GATC : glutamyl-trna(gln) amidotransferase, subunit C HBL : Hospital Barros Luco HER2 : Receptor 2 del factor de crecimiento epidermal. HNF1A : HNF1 homeobox A HOXA9 : homeobox A9 HOXA10 : homeobox A10 HOXA11 : homeobox A11 HOXA13 : homeobox A13 HR : Homology Recombination HSBA : Hospital San Borja Arriarán INC : Instituto Nacional del Cáncer. JJA : Hospital José Joaquín Aguirre JMJD6 : jumonji domain containing 6 KATNB1 : katanin p80 (WD repeat containing) subunit B 1 KLF13 : Kruppel-like factor 13 KNTC1 : kinetochore associated 1 KRT7 : keratin 7 LCE2A : late cornified envelope 2A LCE2B : late cornified envelope 2B LCE2C : late cornified envelope 2C LCE2D : late cornified envelope 2D MACF1 : microtubule-actin crosslinking factor 1 MAP3K8 : mitogen-activated protein kinase kinase kinase 8 MDM2 : MDM2 proto-oncogene, E3 ubiquitin protein ligase x
16 MPHOSPH9 : M-phase phosphoprotein 9 MYC : v-myc avian myelocytomatosis viral oncogene homolog NCOR2 : nuclear receptor corepressor 2 NHEJ : non-homologous end joining NISCH : nischarin NOTCH3 : notch 3 NUDC : nudc nuclear distribution protein P2RX7 : purinergic receptor P2X, ligand-gated ion channel, 7 PALB2 : Partner and Localizer of BRCA2 PARP1 : poly (ADP-ribose) polymerase 1 PEBP1 : phosphatidylethanolamine binding protein 1 PDE4DIP : phosphodiesterase 4D interacting protein PI3K : phosphatidylinositol-4,5-bisphosphate 3-kinase PIWIL1 : piwi-like RNA-mediated gene silencing 1 PKN1 : protein kinase N1 PLA2G1B : hospholipase A2, group IB (pancreas) POLE : polymerase (DNA directed), epsilon, catalytic subunit PR : receptor de progesterona PSMD9 : proteasome (macropain) 26S subunit, non-atpase, 9 PTPN11 : protein tyrosine phosphatase, non-receptor type 11 PUC : Hospital Clínico de la Universidad Católica de Chile RAB20 : RAB20, member RAS oncogene family RAD17 : RAD17 homolog (S. pombe) RAD50 : RAD50 homolog (S. cerevisiae) RAN : RAN, member RAS oncogene family RASGEF1C : RasGEF domain family, member 1C RASGRP1 : RAS guanyl releasing protein 1 (calcium and DAGregulated) RB1 : retinoblastoma 1 RBM14 : RNA binding motif protein 14 RBM4 : RNA binding motif protein 4 RE : receptor de estrógeno RUVBL1 : RuvB-like AAA ATPase 1 SCARB1 : scavenger receptor class B, member 1 SEBOX : SEBOX homeobox SEC22B : EC22 vesicle trafficking protein homolog B (S. cerevisiae) (gene/pseudogene) SIRT4 : sirtuin 4 SPRR1A : small proline-rich protein 1A SPRR1B : small proline-rich protein 1B SPRR2B : small proline-rich protein 2B SPRR3 : small proline-rich protein 3 SRC : SRC proto-oncogene, non-receptor tyrosine kinase SSA : single strand annealing xi
17 TAOK3 : TAO kinase 3 TARBP2 : TAR (HIV-1) RNA binding protein 2 TBX5 : T-box 5 THBS1 : thrombospondin 1 TNBC : Triple Negative Breast Cancer TP53 : tumor protein p53 U2AF1 : U2 small nuclear RNA auxiliary factor 1 UBC : ubiquitin C VTN : vitronectin WDTC1 : WD and tetratricopeptide repeats 1 ZEB1 : zinc finger E-box binding homeobox 1 ZKSCAN3 : zinc finger with KRAB and SCAN domains 3 ZKSCAN4 : zinc finger with KRAB and SCAN domains 4 ZNF160 : zinc finger protein 160 ZNF187 : zinc finger and SCAN domain containing 26 ZNF192 : zinc finger with KRAB and SCAN domains 8 ZNF193 : zinc finger and SCAN domain containing 9 ZNF217 : zinc finger protein 217 ZNF28 : zinc finger protein 28 ZNF323 : zinc finger and SCAN domain containing 31 ZNF415 : zinc finger protein 415 ZNF611 : zinc finger protein 611 ZNF83 : zinc finger protein 83 ZNRD1 : zinc ribbon domain containing 1 ZSCAN16 : zinc finger and SCAN domain containing 16 xii
18 RESUMEN El cáncer de mama triple negativo (TNBC) definido por la ausencia de expresión del receptor de estrógeno, del receptor de progesterona y del receptor 2 del factor de crecimiento epidermal (HER2), representa un grupo agresivo, para el cual no existe hasta ahora una terapia dirigida. Los tumores TNBC frecuentemente se observan en mujeres portadoras de mutación en el gen BRCA1 en la línea germinal y en aproximadamente el 30% de las mujeres no portadoras de mutación, cuyos tumores presentan pérdida de expresión de la proteína BRCA1, sugiriendo que BRCA1 puede ser relevante en el desarrollo de los tumores TNBC. Además, aproximadamente un 70% de los tumores TNBC se agrupan en el subtipo molecular tipo Basal, constituyendo un subgrupo que comparte un perfil de expresión génica similar, con una respuesta clínica diferente. Conocer los genes involucrados en la biología de este tipo de cáncer de mama es relevante para su tratamiento. A este respecto, entre las alteraciones observadas con mayor frecuencia en cáncer encontramos deleciones cromosómicas, conteniendo genes supresores de tumor, y ganancias o amplificaciones involucrando oncogenes. Por lo tanto, identificar regiones genómicas afectadas en su número de copias permite identificar genes o vías de señalización, que pueden corresponder a potenciales blancos de terapia para el tratamiento de estos pacientes. Dado estos antecedentes, se propuso que los tumores de cáncer de mama triple negativo de tipo Basal con pérdida de expresión de la proteína BRCA1, comparten un patrón similar de ganancias y deleciones genómicas. Se estudiaron 48 tumores TNBC por inmunohistoquímica para evaluar la expresión de las citoqueratinas CK5 y CK14, EGFR y la proteína BRCA1. Para definir el subgrupo TNBC/tipo Basal se usó la expresión de las citoqueratinas y EGFR. Además, mediante la técnica de hibridación genómica comparativa en microarreglos de DNA (array-cgh) se identificaron las regiones genómicas de ganancias y deleciones comunes entre los casos TNBC. El análisis de la expresión de las citoqueratinas (CK5 y CK14) y EGFR reveló que un 77% (37/48) de los tumores TNBC se clasifican en el subtipo tipo Basal, principalmente definidos por la expresión de la CK5. Por otra parte, la evaluación de la inmunotinción de BRCA1 en los casos TNBC, xiii
19 mostraron intensidades de tinción variables en el núcleo y expresión en el citoplasma de las células tumorales. A diferencia de los tejidos de mama normal, cuya expresión fue observada en el núcleo de las células epiteliales que forman los ductos y lóbulos. A este respecto, la expresión predominantemente nuclear de BRCA1 se condice con su participación en la reparación del DNA por quiebre en la doble hebra, por lo tanto, su expresión en esta ubicación celular es requerida para su normal funcionamiento. Entonces, debido a los diferentes patrones de expresión y localización de BRCA1 y con el fin de separar los tumores TNBC con ausente o reducida expresión nuclear de BRCA1 que den cuenta de una pérdida de función de BRCA1, los tumores TNBC fueron clasificados en tres subgrupos. Por consiguiente, un 20,8% (10/48) de los tumores TNBC mostraron expresión solo nuclear, con intensidades de tinción similares a los tejidos de mama normal. Un segundo subgrupo se conformó por un 47,9% (23/48) de los casos que revelaron expresión citoplasmática de BRCA1 con o sin expresión nuclear. Finalmente, un 31,3% (15/48) de los tumores TNBC mostraron ausente o reducida expresión de la proteína BRCA1 en el núcleo, sin expresión citoplasmática. Este último subgrupo, principalmente TNBC/tipo Basal representan el grupo central de la hipótesis propuesta en este estudio y fue denominado TNBC/tipo Basal con pérdida de expresión nuclear de BRCA1. Estos resultados sugieren que la pérdida de expresión de BRCA1 o su alterada localización pueden ser relevantes para la progresión del tumor TNBC. El análisis del genoma de los tumores TNBC por array-cgh reveló diferentes alteraciones genómicas recurrentes entre ellos, siendo más frecuente las regiones genómicas de ganancia. El perfil de cambios en el número de copias del DNA fue similar entre los diferentes subgrupos de tumores agrupados por la expresión y localización de BRCA1. Sin embargo, el subgrupo con expresión citoplasmática de BRCA1, reveló una baja frecuencia de cambios en el número de copias del DNA entre ellos, en comparación con los otros dos subgrupos de tumores. Más aún, alteraciones genómicas significativamente diferentes entre los tres subgrupos de tumores con diferente expresión/localización de BRCA1 fueron identificadas. Por su parte, el agrupamiento jerárquico no supervisado basado en xiv
20 7.293 genes comprendidos en las regiones genómicas de ganancia y deleción de los tumores TNBC mostró una clara separación de los tumores con expresión nuclear/citoplasmática de BRCA1 de los otros dos subgrupos de tumores. A este respecto, se encontró que el 70% (16/23) de los tumores con expresión nuclear y citoplasmática de BRCA1 se agruparon en la rama con un bajo número de genes con alteración en su número de copias. En cambio, la rama con mayor número de genes con alteración genómica concentró un 82% de los casos con expresión nuclear y un 73% de los tumores TNBC/tipo Basal con pérdida de expresión nuclear de BRCA1. Sin embargo, para estos dos subgrupos de tumores TNBC se identificaron regiones genómicas de ganancia y deleción estadísticamente significativas. El análisis de la ontología de los genes contenidos en las regiones genómicas de ganancia y deleción reveló procesos biológicos y vías de señalización específicas afectadas en los subgrupos de tumores TNBC agrupados por la expresión y localización de BRCA1. Tumores con expresión nuclear de BRCA1, mostraron ganancia de una región genómica que contiene mtor, un blanco de terapia de la vía de señalización PI3K/AKT/mTOR. Por su parte, los tumores con TNBC/tipo Basal con pérdida de expresión nuclear de BRCA1 mostraron regiones de ganancia genómica que contienen genes involucrados en la transducción de la proteína RAS. Así se identificó RASGRP1 que activa la proteína RAS, revelando que esta vía de señalización puede ser relevante en el tratamiento de paciente con deficiencia de BRCA1. Los resultados sugieren que el análisis de la expresión de la proteína BRCA1, particularmente en el núcleo, permite separar los tumores TNBC en dos subgrupos, dado que específicas alteraciones genómicas revelan una progresión diferente de estos tumores. Además, los tumores TNBC/tipo Basal con pérdida de expresión de BRCA1 con alta inestabilidad genómica y un patrón similar de ganancias y pérdidas genómicas sugiere que pueden ser seleccionados para el tratamiento con inhibidores de PARP1 y pueden tener una mayor sensibilidad al tratamiento con drogas que inducen daño, tales como las drogas de platino. xv
21 ABSTRACT Triple-negative breast cancer (TNBC), defined by the absence of expression of estrogen and progesterone receptors and epidermal growth factor receptor 2 (HER2), represent an aggressive group of tumors lacking directed therapy. In this regard, TNBC tumors are frequently observed in women carrying a germline mutation in the BRCA1 gene and in approximately 30% of non-carrier women with loss of expression of BRCA1 protein in their tumors, suggesting BRCA1 could be relevant in TNBC tumor development and/or progression. In addition, about 70% of TNBC tumors belong to the molecular subtype basal-like, described by gene expression microarray, constituting a biological and clinical distinct subgroup. A knowledge of the genes involved in this type of cancer may have relevance for treatment. In this regard, the most frequently identified alterations in cancer are chromosomal deletions containing tumor suppressor genes and chromosomal gains involving oncogenes. Therefore, identifying regions affected by copy number alterations may help identify genes in molecular or signaling pathways that could be potential treatment targets. To this end we proposed that basal-like triple negative breast tumors with loss of expression of BRCA1 protein share a similar pattern of genomic deletions and gains. We studied 48 triple-negative breast tumors by immunohistochemistry for cytokeratins CK5 and CK14, EGFR and BRCA1. The basal-like subtype was assessed in TNBC tumors by the expression of cytokeratins 5, 14 and EGFR. Using microarray-based comparative genomic hybridization (array- CGH), we characterized common gains and losses between tumors. Our analysis revealed that 77% (37/48) of triple-negative tumors were defined as basal-like, mainly by the expression of CK5. On the other hand, the evaluation of immunostaining of BRCA1 in the tumor cells from TNBC cases showed variable staining intensities between the nucleus and cytoplasm. This contrasted with normal breast tissue where BRCA1 expression was observed in the nucleus of the ductal and lobular epithelial cells. In this respect, the predominantly nuclear expression of BRCA1 is consistent with its participation in the double-strand DNA break repair; thus its expression xvi
22 in this cell location is required for normal function. Consequently, due to the different patterns of expression and localization of the BRCA1 protein and in order to separate the TNBC tumors based on absence or reduced nuclear expression of BRCA1 (indicating a loss of function of BRCA1), TNBC tumors were classified into three subgroups. Subgroup 1 contained 20.8% (10/48) of tumors TNBC showing only nuclear expression of BRCA1, with intensities staining similar to normal breast tissues. A second subgroup constituting 47.9% (23/48) of cases possessed cytoplasmic expression of BRCA1 with/without nuclear expression. Finally, 31.3% (15/48) of tumors showed absent or reduced expression of BRCA1 protein in the nucleus, without cytoplasmic expression. The latter subgroup was mainly comprised of "TNBC/Basal-like" tumors, which corresponded to the central hypothesis of this study. Our results suggest that the loss of expression of BRCA1 or its altered localization may be relevant for the TNBC tumor progression. Analysis of genomic alterations by array-cgh revealed different recurrent genomic alterations in TNBC tumors with a gain of genomic regions being more frequent. The profile of copy number alterations was similar between subgroups of tumors with different expression and localization of BRCA1. However, the frequency of these genomic alterations was lower in the group with cytoplasmic expression than in the other groups. Moreover, we found copy number alterations significantly different among subgroups of tumors with different expression/localization of BRCA1. Unsupervised hierarchical clustering of TNBC using 7,293 genes comprised in copy number gains and losses of TNBC tumors showed a clear separation of the tumors, regarding the expression and localization of BRCA1 protein. In this respect, we found that 70% (16/23) of tumors with nuclear and cytoplasmic expression of BRCA1 grouped in the branch of genes with low numbers of changes in their copy number. In contrast, the branch with the high number of genes altered concentrated 82% of tumors with nuclear expression of BRCA1 tumors and 73% of tumors with "TNBC/Basal-like with loss of nuclear expression of BRCA1." However, these subgroups of tumors have significantly distinct genomic regions of gain and deletion. xvii
23 Ontology analysis of the genes contained in these regions revealed specific biological processes and signaling pathways affected in the three tumor subgroups. In cases with "nuclear expression of BRCA1" there was a gain of genomic regions containing mtor, a target for therapy of the PI3K/AKT/mTOR signaling pathway. In contrast, in the tumors "TNBC/Basal-like with loss nuclear expression of BRCA1 mainly genes related to epithelial cell differentiation and RAS protein transduction were identified. In particular, the identification of the gene RASGRP1 that activates RAS, suggests that this signaling pathway may be relevant in the treatment of BRCA1 deficient patients. Clinical progression suggest the potential separation of TNBC tumors into two groups based on specific genomic alterations in the presence of BRCA1 nuclear expression. In addition, "TNBC/Basal-like with loss nuclear expression of BRCA1 tumors with a high genomic instability and similar patterns of gain and loss within genomic region suggests the potential for personalized use of PARP inhibitors and a greater sensitivity to DNA-damaging agents such as platinum. xviii
24 INTRODUCCION 1. El cáncer de mama. El cáncer de mama, es una enfermedad frecuente a nivel mundial y en Chile es la primera causa de muerte por cáncer en mujeres, con una tasa de mortalidad estandarizada de 15,5 x mujeres (DEIS., ). El fenotipo de esta enfermedad compleja resulta de la acción combinada de factores genéticos y ambientales. Se presenta de forma esporádica o hereditaria. Se ha estimado que la forma hereditaria definida por la historia familiar de cáncer de mama y/u ovario en parientes de primer grado representa cerca de un 10% de los casos totales de cáncer de mama (National Comprehensive Cancer Network (NCCN)). El cáncer de mama y/u ovario hereditario a diferencia del cáncer de mama esporádico se puede presentar a una edad muy temprana ( 40 años de edad), y con alta prevalencia de cáncer de mama bilateral (Loman y cols., 2001). Con el desarrollo de nuevas tecnologías, el estudio en cáncer de mama ha abierto nuevos horizontes a nivel inmunofenotípico, genómico y molecular que han permitido vislumbrar similitudes y diferencias entre los tumores de pacientes con cáncer de mama hereditario y los esporádico (van der Groep y cols., 2006; Honrado y cols., 2006). 2. Genes de alto riesgo al cáncer de mama hereditario (BRCA1 y BRCA2) Dos genes de alto riesgo al cáncer de mama han sido descubiertos hasta la fecha. Estos son: BRCA1, localizado en el cromosoma 17q21 (Miki y cols., 1994) y BRCA2, ubicado en el cromosoma 13q12 (Wooster y cols., 1995). Mutaciones en la línea germinal en uno de estos dos genes confieren un alto riesgo de desarrollar cáncer de mama y/u ovario hereditario (Narod y cols., 2004; King., 2014). La proporción de familias con mutación en uno de estos genes BRCA1 o BRCA2 varía de acuerdo a la población analizada y los criterios de selección de las pacientes 1
25 (Fackenthal y cols., 2007; Díez y cols., 2003; Narod y cols., 2004). En Chile, un 20% de las familias presentan mutación en uno de estos dos genes (Gallardo y cols., 2006), similar a la población española que asciende a un 25% de los casos de cáncer de mama (Diez y cols., 2003). Sin embargo, aún se desconoce la causa genética para el 80% de las familias restantes. Nuestro estudio genético en Chile, sumado a los realizados en otras poblaciones del mundo, plantean la existencia de otro u otros genes como factores de genéticos de alto riesgo al cáncer de mama hereditario (Diez y cols., 2003; Gallardo y cols., 2006; King., 2014). A este respecto, existen mutaciones asociadas a diversos síndromes como: Li Fraumeni, Cowden, y otros en los cuales se presentan casos de cáncer de mama. Para estos síndromes se han descrito mutaciones en los genes TP53 y PTEN respectivamente (Birch y cols., 1994; Liaw y cols., 1997). Por otra parte, mutaciones en los genes ATM, BRIP1, PALB2, y la variante alélica 1100delC del gen CHEK2, son consideradas de moderada penetrancia, y confieren un riesgo menor a cáncer de mama (Meijers- Heijboer y cols., 2002; Renwick y cols., 2006; Seal y cols., 2006; Rahman y cols., 2007). Nuestro grupo realizó el análisis de estos últimos genes en familias chilenas con cáncer de mama y/u ovario hereditario no portadoras de mutación en BRCA1 o BRCA2, y no se identificaron mutaciones en estos genes que afecten la función de sus proteínas respectivas o incluso que segreguen con la enfermedad en la familia (Tapia T y cols., 2008; y manuscrito en preparación). Esto demuestra la heterogeneidad genética del cáncer de mama hereditario y la importancia de encontrar el gen o los genes involucrados en las génesis de esta enfermedad, responsables de su progresión (Honrado y cols 2006; Couch y cols., 2014). 2
26 3. Clasificación del cáncer de mama, tanto de esporádico y hereditario, en relación a la histología e inmunofenotipo. El cáncer de mama, no sólo es una enfermedad heterogénea a nivel genético, también lo es fenotípica, clínica y molecularmente, reflejándose directamente en el diagnóstico, el pronóstico, la respuesta clínica y el tratamiento otorgado a las pacientes. (Sorlie y cols., 2004). Las principales interrogantes de estos diversos estudios apuntan a descubrir cuáles son los genes que originan el proceso neoplásico y cuales son aquellos que sustentan su desarrollo, la invasión de tejidos y la metástasis. La mayoría de los estudios están enfocados en la identificación de una o varias moléculas que puedan ser blancos de un mejor o un nuevo tratamiento para esta enfermedad (Guedj y cols., 2012; The Cancer Genome Atlas Network., 2012; Vogelstein y cols., 2013). A nivel fenotípico, los tumores de mama son clasificados por los anátomo-patólogos, en base a características morfológicas descriptivas del tumor y de sus células neoplásicas (Weigelt y cols., 2009). La histología del carcinoma es determinada por la localización de las células tumorales, y pueden ser ductal o lobulillar, invasivo o in situ, además permite determinar el grado del tumor (estado de diferenciación de las células tumorales), la presencia de invasión linfocitaria, la presencia de necrosis, mitosis y en el caso de los linfonodos la existencia de metástasis (Weigelt y cols., 2009). La histología más recurrente corresponde al carcinoma ductal invasor, con aproximadamente un 70% del cáncer de mama (Tavassoli y cols., 2003). En este grupo histológico se concentran los tipos histológicos denominados de: tipo no especial y aquellos de tipo especial que incluye, el tipo metaplásico, el tipo medular, el tipo mucinoso y el tipo papilar entre otros, siendo más frecuente los carcinomas ductal invasor de tipo no especial (Tavassoli y cols., 2003). Los patrones definidos por la morfología del tumor y las células neoplásicas vistas al 3
27 microscopio son utilizados para la clasificación actual de los tumores en la mayoría de los laboratorios de anatomía-patológica (Tavassoli y cols., 2003; Weigelt y cols., 2009). Una segunda clasificación de los tumores de mama, se basa en el análisis de la expresión mediante inmunohistoquímica del receptor de estrógeno, el receptor de progesterona y la sobreexpresión del receptor 2 del factor de crecimiento epidermal (HER2), y se relaciona con los comienzos de la terapia dirigida del cáncer de mama (Carey y cols., 2010). Estos marcadores proteicos, de gran importancia en la clínica, permiten determinar el pronóstico y escoger la estrategia de tratamiento a las pacientes con esta enfermedad. Así, pacientes con tumores de mama que expresan el receptor de estrógeno, pueden tratarse con anti-estrógenos, como el tamoxifeno, cuyo metabolito activo, denominado endoxifeno, compite con el estrógeno por los sitios de unión de ligando del receptor (Hoskins y cols., 2009). Otra terapia, preferente en mujeres postmenopáusicas, consiste en el uso de inhibidores de la enzima aromatasa, que previene la síntesis de estrógeno en los tejidos periféricos, incluida la mama (Di Cosimo y cols., 2010). Estos tumores representan aproximadamente el 70% de los casos de cáncer de mama y son de buen pronóstico para las pacientes (Tavassoli y cols., 2003). Por su parte, la expresión del receptor de progesterona, es un indicador de buen pronóstico para los tratamientos previamente mencionados, ya que revela la funcionalidad del receptor de estrógeno y aumenta la respuesta al tamoxifeno y a los inhibidores de aromatasa (Stendahl y cols., 2006; Dowsett y cols., 2005; Purdie y cols., 2014). Otro grupo de tumores con un tratamiento dirigido, corresponde a aquellos que expresan el receptor 2 del factor de crecimiento epidermal (HER2) en las membranas de las células tumorales. Este grupo representa aproximadamente entre un 15-30% de los casos de cáncer de mama, son de mal pronóstico, agresivos, e indican una disminución de la sobrevida de las pacientes (Tavassoli y cols., 2003). Sin embargo, estos parámetros se ven mejorados por el tratamiento con un 4
28 anticuerpo monoclonal denominado, Traztuzumab, el cual se une al dominio extracelular de HER2, impidiendo la homo o heterodimerización de este receptor con otros miembros de la familia de los receptores del factor de crecimiento epidermal, y por lo tanto su funcionalidad. (Di Cosimo y cols., 2010). Finalmente, un tercer grupo se origina por exclusión, ya que carece de la expresión de estos tres receptores y no pueden ser tratados con los tratamientos dirigidos mencionados previamente. Así, surgen los tumores de cáncer de mama con fenotipo triple negativo, como un grupo particular de cáncer de mama (Carey y cols., 2010). 4. Definición de los tumores de mama con fenotipo triple negativo Los tumores de mama triple negativo se definen por la ausencia de expresión del receptor de estrógeno (RE), del receptor de progesterona (RP) y del receptor 2 del factor de crecimiento epidermal (HER2), mediante inmunohistoquímica. (Carey y cols., 2010; Linn y van 't Veer., 2009; Metzger-Filho y cols., 2012). Este grupo particular de tumores representa entre un 10-24% de los casos de cáncer de mama (Bauer y cols., 2007; Carey cols., 2010). Su mayor prevalencia se ha identificado en mujeres premenopáusicas de origen africano y en africanas-americanas (Carey y cols. 2010; Linn y Van 't Veer., 2009). Los tumores de mama triple negativo se caracterizan por su alto grado histológico (grado III), agresivos, de mal pronóstico, de ocurrencia a edad temprana y de elevado riesgo de recurrencia en comparación con otros grupos de tumores de mama (Cleator y cols., 2007; Reis-Filho y cols., 2008; Irvin y cols., 2008). La mayoría de estos tumores corresponden a carcinoma ductal invasor, con elevado conteo mitótico y presencia de necrosis centrales. Pocas pacientes con este tipo de tumor presentan una buena respuesta a la quimioterapia con antraciclinas y taxanos (Carey y cols., 2010), careciendo de un tratamiento efectivo y no cuentan con un tratamiento dirigido. Por esto, 5
29 es relevante conocer los genes y las vías moleculares que participan en el inicio y progresión de este tipo de tumor, con el fin de ofrecer a las pacientes un mejor tratamiento y pronóstico. 5. Clasificación molecular de los tumores de mama, basada en los perfiles de expresión génica. El desarrollo de la técnica de microarreglos de DNA, permitió clasificar los tumores de mama según sus perfiles de expresión génica. Perou y cols., (2000) a partir del estudio de 40 pacientes con cáncer de mama esporádico, realizaron un agrupamiento jerárquico de los tumores, en base a la similaridad de sus perfiles de expresión génica, determinados por microarreglos de expresión del mrna (Perou y cols., 2000). Esto, llevó a identificar 6 subgrupos de tumores de mama con diferentes perfiles de expresión génica, pero similares entre los tumores del mismo subgrupo (figura 1) (Perou y cols., 2000, Sorlie y cols., 2001). En la figura 1, se observa que las dos principales cladas se diferencian por los niveles de expresión del receptor de estrógeno. Los tumores que expresan este receptor se subdividen en tres subgrupos, denominados: Luminal A, con alta expresión del receptor de estrógeno, y de los factores de transcripción GATA3, XBP1 y HNF3A (Perou y cols., 2000; Sorlie y cols., 2001). Por su parte, los tumores con moderada expresión del receptor de estrógeno fueron llamados luminal B y C. Estos tres subgrupos expresan genes específicos de las células epiteliales luminales, siendo el luminal A de mejor pronóstico que los luminal B/C (figura 2). Por su parte, los tumores de la clada con ausente expresión del receptor de estrógeno se agruparon en los subgrupos denominados: HER2+, tipo mama normal y el tipo Basal. El subgrupo HER2+, se caracteriza por una alta expresión del receptor 2 del factor de crecimiento epidermal (HER2). El subgrupo tipo mama normal presento alta expresión de genes que se expresan en el tejido adiposo y en otros tipos celulares no epiteliales, como las células estromales. Además, expresan genes relacionados a las células basal/mioepitelial y baja 6
30 expresión de genes epiteliales luminales (Perou y cols., 2000; Sorlie y cols., 2001). Por último, el subgrupo tipo Basal fue caracterizado por la alta expresión de las citoqueratinas 5 (CK5) y 17 (CK17) que se expresan normalmente en el epitelio basal/mioepitelial de la glándula mamaria normal, y expresaron altos niveles de laminina, P-cadherina, el receptor de integrina (α6β4), vimentina y el receptor del factor de crecimiento epidermal (EGFR) (Sorlie y cols., 2001). Este último expresado en aproximadamente el 60% de los tumores tipo Basal (Schneider y cols., 2008, Miyoshi y cols., 2008). Esta clasificación molecular de los tumores de mama ha sido confirmada por otros grupos de investigación en diferentes poblaciones de pacientes con cáncer de mama (Sorlie y cols., 2003; Bertucci y cols., 2008; Di Cosimo y cols., 2010). La importancia de esta clasificación molecular en la clínica se revela en el trabajo de Sorlie y cols., (2001) en el cual se demuestra que esta clasificación molecular, basada en los perfiles de expresión génica, se correlacionan con la sobrevida de los pacientes con esta enfermedad (Sorlie y cols., 2001). El análisis de la sobrevida total y de la sobrevida libre de recurrencia de las pacientes con tumores de mama, según la clasificación molecular, revela que los subtipos tipo Basal y HER2+ se asocian con un corto tiempo de sobrevida total y sobrevida libre de recurrencia en comparación con los tumores del subtipo Luminal A y tipo mama normal. Por su parte, los tumores agrupados en los subtipos luminal B y C presentan un menor tiempo de sobrevida libre de recurrencia que los tumores del subtipo Luminal A (figura 2) (Sorlie y cols., 2001). Esto sugiere que existe un grupo de genes que se expresan en los tumores luminal B y C similares a los subtipos tipo Basal y HER2+ asociados con una pobre respuesta clínica de las pacientes. (Sorlie y cols., 2001; Sorlie., 2004). 7
 Perfil de expresión génica de los 78 carcinomas, 3 tumores benignos y 4 tejidos de mama normal, agrupados en 6 subtipos diferentes, basándose en una lista intrínseca de 500 genes.")
, Luminal subtype C (tipo luminal C), Luminal subtype A (tipo luminal A). B) Diagrama del agrupamiento de los genes específicos de cada subgrupo.")
31 A) RE(-) RE(+) B) Figura 1: Agrupamiento jerárquico de los tumores de cáncer de mama en 6 subtipos diferentes. A) Perfil de expresión génica de los 78 carcinomas, 3 tumores benignos y 4 tejidos de mama normal, agrupados en 6 subtipos diferentes, basándose en una lista intrínseca de 500 genes. En esta figura se muestra que los tumores de mama (carcinomas) fueron divididos en 6 subtipos principales denominados Basal-like (tipo Basal), ERBB2+ (HER2+), Normal breast-like (tipo mama normal), Luminal subtype B (tipo luminal B), Luminal subtype C (tipo luminal C), Luminal subtype A (tipo luminal A). B) Diagrama del agrupamiento de los genes específicos de cada subgrupo. Se indica en C: Genes del amplicon HER2+; en D: genes nuevos desconocidos; en E: grupo de genes enriquecidos del epitelio basal; en F: Grupos de genes de la mama normal; en G: grupo de genes del epitelio luminal, conteniendo el receptor de estrógeno (RE). Esta figura fue extraída del artículo publicado por Sorlie y cols.,
32 Probabilidad Probabilidad A B Sobrevida (Meses) Sobrevida libre de recurrencia X Censorados, Lum A, Lum B+C, Tipo NorB, Tipo Basal, HER2+ Figura 2: Curvas de Kaplan-Meier de la sobrevida total y de la sobrevida libre de recurrencia de 49 pacientes con cáncer de mama, basada en la clasificación de los diferentes perfiles de expresión génica. (A) Sobrevida total y B) Sobrevida libre de recurrencia para los 5 subtipos moleculares definidos como Lum A, Luminal A; Lum B+C, Luminal B y Luminal C que fueron considerados como un solo grupo; Tipo NorB, tipo mama norma; tipo Basal; HER2+. En la curva de Kaplan-Meier, cada subtipo molecular está representado con un color respectivo, como se muestra en la parte inferior de la figura. Las curvas en la figura muestran que los tumores tipo Basal, HER2+ y el conjunto de los tumores Luminal B y C tienen un menor tiempo de sobrevida total y libre de recurrencia en comparación con los subtipos moleculares restantes. (Figura extraída y modificada desde la publicación de Sorlie y cols., 2001). 9
33 6. Estructura de la glándula mamaria, y las células epiteliales que la forman. La glándula mamaria es un órgano dinámico y estructurado que cambia con la edad, la pubertad y el embarazo (Gusterson y Stein, 2012). Consiste de una estructura ramificada llamada túbulo-alveolar, conformada por ductos y lobulillos que contienen los grupos de acinos secretores (figura 3). Los ductos y lobulillos, están hechos de un epitelio estratificado, de dos tipos de células epiteliales. Las células luminales, que revisten el lumen interno responden al estrógeno y corresponden a las productoras de la leche. Las células basal/mioepitelial (término usado intercambiablemente para describir la localización basal de estas células epiteliales con función contráctil) son localizadas bajo las células luminales y en contacto con la membrana basal. Estas células proveen la fuerza contráctil para expeler la leche durante la lactancia (Gusterson y Stein, 2012). Se ha descrito que ambos tipo celulares se originan a partir de una célula madre común, la cual, carece de expresión del receptor de estrógeno (Sleeman y cols., 2007; Molyneux y cols., 2011). Más aún, las células luminales progenitoras mayoritariamente son negativas para el receptor de estrógeno mientras que las células luminales con expresión de este receptor en gran parte corresponden a células diferenciadas (Sleeman y cols., 2007). Tanto las células epiteliales de la mama, como de otros tejidos estratificados o simples, tienen expresión diferencial de citoqueratinas tipo I y tipo II. De esta manera, las células luminales expresan las citoqueratinas CK7 y CK8 del tipo I y las CK18 y CK19 que son del tipo II. Por otra parte, las células basales (y/o mioepiteliales) expresan las citoqueratinas CK5 y CK17 que son del tipo II y la CK14, del tipo I (figura 3) (Moll y cols., 2008). Esta expresión diferencial de las proteínas de los filamentos intermedios de las células de la mama normal, han sido observadas posterior al proceso neoplásico que sufren las células en el proceso de transformación (Moll y cols., 1982; Taylor-Papadimitriou 10
34 y cols., 1989). Por este motivo, se sugiere que la expresión de estas citoqueratinas permitiría inferir el origen celular del tumor. 7. Identificación de los tumores de mama tipo Basal mediante inmunohistoquímica. Si bien, la definición de los tumores de mama tipo Basal fue determinada por su perfil de expresión génica, esta se simplificó a una metodología más simple, rutinaria en clínica y menos costosa, la técnica de inmunohistoquímica. Nielsen y cols., (2004) evaluaron por inmunohistoquímica la expresión del receptor de estrógeno, del receptor 2 del factor de crecimiento epidermal (HER2), de las citoqueratinas 5 y 6 (CK5/6) y del receptor de factor de crecimiento epidermal (EGFR) en 21 tumores de mama tipo Basal (previamente clasificados por microarreglos de expresión). Estos autores, encontraron que un 76% (16/21) de los casos tipo Basal presentaron ausente expresión del receptor de estrógeno, de HER2 y expresaron CK5/6 y/o EGFR. Con esta estrategia, de un panel de 4 marcadores proteicos, lograron definir los tumores tipo Basal con un 76% de sensibilidad y un 100% de especificidad (Nielsen y cols., 2004). Así, la evaluación por inmunohistoquímica de estos 4 marcadores proteicos permite determinar robustamente tumores tipo Basal. Por consiguiente, otros autores han utilizado esta estrategia para definir los tumores tipo Basal (Rakha y cols., 2009; Cheang y cols., 2008). Aún más, otros estudios han agregado a este panel de marcadores, la expresión del receptor de progesterona mediante inmunohistoquímica con el fin de identificar tumores de mama tipo Basal (Rakha y cols., 2009; Carey y cols., 2007; Dent y cols., 2007; Bauer y cols., 2007), indicando que una alta proporción de los tumores de mama triple negativo, sumado a la expresión de citoqueratinas y de EGFR, definidos por inmunohistoquímica, se corresponden con la definición original del subgrupo tipo Basal. 11

35 Ducto Ductonormal Lóbulo Lobulillo Célula epitelial basal/ mioepitelial Expresión de CK5/6 Expresión de CK14 Expresión de CK17 Ducto de glándula mamaria Célula epitelial luminal Expresión de CK8 Expresión de CK18 Expresión de CK19 Figura 3: Representación esquemática de la glándula mamaria normal, las estructuras y los tipos de células epiteliales que la forman. Se indican el ducto y el lobulillo que forman la glándula mamaria normal. Además, se muestra un ducto, donde se indican las células basal/mioepitelial (color celeste) y las células epiteliales luminal (blancas). En ambos, tipos celulares se indican las citoqueratinas que se expresan específicamente. De esta manera, las células basal/mioepitelial expresan las citoqueratinas CK5, CK14, CK17. Por su parte, las células luminales expresan las citoqueratinas CK8, CK18, CK19. 12
36 Una reciente forma de identificar los tumores tipo Basal se basa en un perfil de expresión, mediante RT-PCR, de 50 genes denominado PAM50 (Parker y cols., 2009). Esta estrategia, evalúa los niveles de expresión de 50 genes que permiten diferenciar los subgrupos moleculares descritos por Perou y cols., (2000). Estos genes derivan de la lista inicial de Sorlie y cols., (2001) usada para clasificar a los tumores de mama en cada subgrupo (Parker y cols., 2009). La importancia de definir estos subgrupos moleculares, mediante cualquier estrategia, se relaciona con seleccionar tumores de mama que permitan ayudar en mejorar el pronóstico de las pacientes y predecir un mayor beneficio de la quimioterapia (Parker y cols., 2009). 8. El grupo de tumores de mama triple negativo integra tumores del subgrupo tipo Basal Los tumores de mama del subgrupo tipo Basal, descritos por Perou y cols., (2000) no expresan el receptor de estrógeno y el receptor 2 del factor de crecimiento epidermal (HER2) (Perou y cols., 2000; Sorlie y cols., 2001). Estos tumores, por primera vez se describieron en el año 1989 (Taylor-Papadimitriou y cols., 1989), como un grupo de tumores de cáncer de mama que se originan desde la capa basal/mioepitelial del epitelio mamario, ya que usualmente expresan las citoqueratinas basales: CK5, CK14 y CK17 (Gusterson y cols., 2005; Linn y Van 't Veer., 2009), pero sólo en los últimos años han sido extensamente mencionados, por la definición molecular descrita por Perou y cols. (2000). Los tumores tipo Basal, representa aproximadamente el 15% de los casos de cáncer de mama y comparten un perfil de expresión génica similar (Perou y cols., 2000; Sorlie y cols., 2001; Bertucci y cols., 2008). Morfológicamente, se caracterizan por ser pobremente diferenciados (Grado III), con alto índice mitótico, presencia de zonas centrales necróticas e infiltración linfocitaria (Fulford y cols., 2006; Livasy y cols., 2006). Más aún, los tumores tipo metaplásico y tipo medular son más prevalentes entre los tipo Basal que en el resto de los subgrupos moleculares de tumores de mama (Jacquemier y cols., 2005; Livasy y cols., 13
37 2006). Por otra parte, estos tumores frecuentemente portan mutaciones en el gen TP53 (Sorlie y cols., 2001) y expresan en mayores niveles genes relacionados con la proliferación celular, comparado con los otros subgrupos moleculares (Perou y cols., 2000; Sorlie y cols., 2003; Hu y cols., 2006). Estas características morfológicas, inmunofenotípicas y moleculares de los tumores de mama del subgrupo tipo Basal, llevaron a determinar que existe similitud con los tumores de mama triple negativo, definidos por la inmunohistoquímica de tres receptores (Carey y cols., 2010). Asi, un estudio mediante microarreglos de expresión génica de 97 tumores de mama triple negativo, definidos por la ausencia del receptor de estrógeno, de progesterona y de HER2, reveló que un 91% de estos tumores triple negativo fueran clasificados como tumores tipo Basal (Kreike y cols., 2007). Otros estudios realizados en cáncer de mama, a través de microarreglos de expresión génica, han confirmado que los tumores de cáncer de mama triple negativo forman un grupo que se sobrepone con los tumores tipo Basal en un 70-80% de los casos. Así, entre un 20-30% de los tumores de cáncer de mama triple negativo no son definidos en el subgrupo tipo Basal y entre un 20-30% de los tumores tipo Basal, no se corresponden con un fenotipo triple negativo (Livasy y cols., 2006; Kreike y cols., 2007; Bertucci y cols., 2008). Por lo cual, estos dos grupos de tumores de mama no se consideran sinónimos (Rakha y cols., 2007). Pero, la existencia de tumores triple negativo definidos como tipo Basal, ya sea por el perfil de expresión o por inmunohistoquímica, sugiere que corresponden a un grupo homogéneo, que comparten un perfil de expresión génica similar entre ellos (Kreike y cols., 2007; Nielsen y cols., 2004; Rakha y cols., 2009). Aún más, se ha descrito que estos tumores de cáncer de mama triple negativo definidos como tipo Basal expresan los marcadores de proliferación p53, p16 y c-kit (Nielsen y cols., 2004; Rakha y cols., 2009). 14
38 Por otra parte, un estudio en microarreglos de expresión génica de tumores de mama realizado por Prat y col., (2010) describe un agrupamiento jerárquico de tumores de mama que indican la existencia de un nuevo subgrupo llamado tipo bajo-claudina, ubicado en una clada próxima a los tumores de mama tipo Basal. Estos tumores tipo bajo-claudina se caracterizan por una inconsistente expresión de los marcadores basales (CK5 y CK14), además de una ausente expresión del receptor de estrógeno, de progesterona y de HER2, es decir, triple negativo. Este subgrupo molecular está formado por tumores que presentan un enriquecimiento de células con propiedades similares a las células madres y genes relacionados con la transición epiteliomesénquima e interferones (Prat y cols., 2010). Además, en relación a la respuesta clínica, los tumores tipo bajo-claudina presentan un mejor pronóstico que los tumores de mama tipo Basal. Esto indica que los tumores de mama triple negativo, definidos por la inmunohistoquímica de los tres receptores corresponden a un grupo heterogéneo compuesto por al menos dos subgrupos moleculares. 9. Tumores de pacientes portadoras de mutación en el gen BRCA1 se relacionan con el fenotipo triple negativo y el tipo Basal. Otro grupo de tumores de mama corresponde a los tumores de mama provenientes de pacientes con cáncer de mama hereditario. Se ha descrito que los tumores de portadoras de una mutación en la línea germinal del gen BRCA1, usualmente son pobremente diferenciados (grado III), con altos índices de proliferación y alto contenido de infiltrado linfocitario (Lakhani y cols., 1998, Honrado y cols., 2006). Características morfológicas descritas, a su vez, para los grupos de tumores de cáncer de mama triple negativo y los tumores tipo Basal ya sean hereditario o esporádico (Honrado y cols., 2006; Melchor y cols., 2008). Asimismo, mutaciones en el gen TP53 ocurren en mayor frecuencia en los tumores con mutación en BRCA1, que en los casos esporádicos, 15
39 sugiriendo que la pérdida de función de p53 promueve la tumorigenesis en los tumores con pérdida de función de BRCA1, producto de una mutación en el gen (Holstege y cols., 2009). El inmunofenotipo de estos tumores de portadoras de mutación en el gen BRCA1 en un 75% de los casos, presentan ausente expresión del receptor de estrógeno, del receptor de progesterona y del receptor 2 del factor de crecimiento epidermal (HER2) (Lakhani y cols., 2002; Grushko y cols., 2002; Foulkes y cols., 2003; Honrado y cols., 2006; Drost y Jonkers., 2014), revelando un fenotipo triple negativo (Atchley y cols., 2008; Lee y cols., 2011; Reis-Filho y Tutt., 2008). Evidencias tempranas en líneas celulares de cáncer de mama revelaron que BRCA1 inhibe la actividad transcripcional del receptor de estrógeno (Fan y cols., 1999; Xu y cols., 2005). Entre los genes blancos de la activación transcripcional de receptor de estrógeno se encuentra el receptor de progesterona (Horwitz y cols., 1978). Así, BRCA1 puede suprimir el desarrollo del tumor por atenuar la señalización proliferativa activada por el receptor de estrógeno (Fan y cols 2002; Xu y cols., 2005). Sin embargo, frecuentemente los tumores con mutación en BRCA1 tienen ausente expresión del receptor de estrógeno. A este respecto, Hosey y cols., (2007) demostraron que el factor de transcripción OCT1 recluta BRCA1 al promotor del gen ESR1, que codifica el receptor de estrógeno, para activar su transcripción. Por lo tanto, BRCA1 funcional es capaz de modular la expresión del receptor de estrógeno (Hosey y cols., 2007). Por otra parte, el estudio realizado por Sorlie y cols., (2001) mediante microarreglos de expresión génica, mostró que los tumores de mama de portadoras de mutación en BRCA1 se agrupan en la subclase tipo Basal, revelando que ellos comparten un perfil de expresión génica similar (Sorlie y cols., 2003). Además, mediante inmunohistoquímica, se ha descrito que las células tumorales provenientes de portadoras de mutación en BRCA1 expresan los marcadores 16
40 basales CK5/6, CK14, CK17 y el receptor del factor de crecimiento epidermal (EGFR) en mayor proporción que los tumores de portadoras de mutación en el gen BRCA2 y los tumores esporádicos (Rakha y cols., 2009; Foulkes y cols., 2003; Sorlie y cols., 2003; Lakhani y cols., 2005). De este modo, la caracterización de estos marcadores permitió encontrar que cerca de un 70% de los tumores con mutación en el gen BRCA1 se corresponden con el subgrupo tipo Basal (Díaz y cols., 2007; Melchor and Benítez., 2008). Aún más, a nivel molecular, los tumores de portadoras de mutación en BRCA1 frecuentemente portan amplificación de c-myc y alta inestabilidad genómica, al compararlos con otros tumores de mama hereditario (Lakhani y cols., 2002). Estas evidencias morfológicas y moleculares sugieren que existe una relación entre el fenotipo triple negativo, la clasificación tipo Basal y la presencia de mutaciones en el gen BRCA1, revelando una pérdida de función de la proteína BRCA1, en los tumores definidos por el inmunofenotipo y el perfil de expresión génica de los tipo Basal. Si bien, estas evidencias están en el contexto de tumores de pacientes con cáncer de mama hereditario, la pérdida de función de la proteína BRCA1 puede ocurrir mediante otros mecanismos diferentes a una mutación en la línea germinal, que puede ocurrir tanto en tumores de cáncer de mama hereditario sin mutación en el gen BRCA1 como en tumores esporádicos. 10. Pérdida de expresión y mecanismos de pérdida de función de BRCA1. Como se mencionó previamente, la mutación en la línea germinal del gen BRCA1 es un factor de riesgo para el desarrollo del cáncer de mama hereditario. BRCA1 es un gen supresor de tumor, por lo tanto se sugiere que pierde su función celular si ambos alelos han perdido su expresión. La hipótesis del segundo evento propuesta por Knudson (Knudson, 1971) describe la ocurrencia de un primer evento en uno de los alelos de un gen, que puede estar mutado en la línea 17
41 germinal, predisponiendo al cáncer hereditario. En este punto, el individuo heterocigoto no desarrolla cáncer debido a la existencia de una copia con función normal. Sin embargo, luego, ocurre un segundo evento que involucra la inactivación del alelo normal (no mutado) en aquella célula somática, lo cual se denomina pérdida de heterozigocidad (LOH), dando inicio al desarrollo tumoral. La pérdida de función de otros supresores de tumor durante la progresión del cáncer, ocurre de la misma forma, por dos eventos moleculares sucesivos en ambos alelos del gen supresor tumoral. Así, la pérdida de heterozigocidad de BRCA1 en tumores de cáncer de mama hereditario ha sido demostrada en pacientes portadoras de mutaciones en el gen BRCA1 (Merajver y cols., 1995). Frecuentemente, estos tumores presentan pérdida de función del alelo normal, mediante deleción genómica, la cual ha sido estimada que ocurre entre un 50-90% de estos tumores (Merajver y cols, 1995). Otro mecanismo implicado en la pérdida de función de BRCA1, ocurre producto del silenciamiento de la expresión de BRCA1, que corresponde a la metilación del promotor del gen BRCA1, el cual se ha encontrado hipermetilado entre un 42-51% de los casos de cáncer de mama hereditario (Honrado y cols., 2006; Tapia y cols., 2008). Estos diferentes mecanismos conllevan a la pérdida de expresión y consecuentemente pérdida de función de esta proteína en los tumores de pacientes con cáncer de mama hereditario (Honrado y cols., 2006). Por otra parte, en relación a los tumores de mama esporádicos, se ha descrito que un muy bajo porcentaje de estos portan una mutación somática en el gen BRCA1. Pero, recientes estudios en casos de cáncer de mama, mediante la secuenciación de segunda generación, se ha logrado identificar mutaciones somáticas en el gen BRCA1 en una proporción de las células tumorales (The Cancer Genome Atlas Network., 2012). Por otra parte, la inactivación de ambos alelos puede ser causada por otros mecanismos moleculares, como deleción parcial o total del gen y metilación del 18
42 promotor (Merajver y cols, 1995; Catteau y cols, 1999; Esteller y cols, 2000). A este respecto, entre un 20 a un 54% de los tumores de cáncer de mama esporádicos presentan ausente o reducida expresión de la proteína BRCA1, y no portan mutaciones somáticas en este gen (Taylor y cols., 1998; Lambie y cols, 2003, Lee y cols., 1999; Yoshikawa y cols., 1999; Rakha y cols., 2008). A su vez, hipermetilación del promotor de BRCA1 ha sido descrito entre un 11-30% de los tumores con cáncer de mama esporádico (Catteau y cols., 1999; Wei y cols., 2005). De esta manera la pérdida de función de BRCA1 está implicada en el cáncer de mama, ya sea éste hereditario o esporádico. En relación, al grupo de tumores de mama triple negativo, se ha descrito que entre un 25-50% de los tumores triple negativo, esporádico o hereditario, presentan hipermetilación del promotor de BRCA1 (Lips y cols., 2013; Xu y cols., 2013). Además, se ha observado que un 30% de los tumores de cáncer de mama triple negativo esporádico tienen ausencia de expresión de la proteína BRCA1. Por su parte, en los tumores clasificados como tipo Basal, la metilación del promotor de BRCA1 se ha identificado en un 30% de estos casos (Lips y cols., 2013) Así, este mecanismo es relevante para reducir la expresión de BRCA1 en los tumores de mama clasificados como triple negativo o tumores tipo Basal. Otros mecanismos implicados en reducir la expresión de BRCA1 y llevar a una pérdida de función de la proteína corresponde a la regulación transcripcional de BRCA1. A este respecto, en tumores de mama tipo Basal se ha propuesto que el aumento de expresión de ID4, un represor transcripcional, disminuye la expresión de BRCA1, sugiriendo un potencial mecanismo para la disminución de expresión de BRCA1 (Beger y cols., 2001; Wen y cols., 2012). Asimismo, se han descrito algunos micrornas, como mir-146a y mir-638, que regulan la expresión de BRCA1 y 19
43 que están asociados al fenotipo molecular tipo Basal. (Chang and Sharan 2012; Shen y cols., 2008). 11. Localización de la proteína BRCA1 en los tumores de mama. En las células epiteliales mamarias normales, la proteína BRCA1 se localiza principalmente en el núcleo, donde realiza una de sus principales funciones celulares que corresponde a la reparación del DNA. (Chen y cols., 1995; Chen y cols., 1996; Scully y cols., 1996; Thomas y cols., 1996). Sin embargo, en las células de tumores de mama, ésta se ha encontrado también en el núcleo y/o citoplasma. (Wilson y cols., 1999; Rakha y cols., 2008). Por consiguiente, además de una reducida expresión de la proteína BRCA1 en el núcleo de las células tumorales, diversos autores han descrito una alterada localización subcelular de esta proteína. (Chen 1995; Coene y cols., 1997; Wilson y cols., 1999; Lee 1999; Yoshikawa y cols., 1999; Rakha y cols., 2008). Esto sugiere, que los tumores con expresión de BRCA1 en el citoplasma, podría inferir una pérdida de función de BRCA1 en estos tumores. Así, la ausencia de BRCA1 en el núcleo y su translocación al citoplasma puede ser una estrategia terapéutica potencial (Drost y Jonkers. 2014). Se ha sugerido que la expresión citoplasmática de BRCA1 puede ser producto de mutaciones somáticas que perjudican la translocación de esta proteína al núcleo (Rodríguez y cols., 2004). Por otra parte, distintas isoformas de BRCA1, producto de un splicing alternativo de BRCA1, pueden localizarse en el citoplasma de las células tumorales. A este respecto, se ha descrito que la isoforma BRCA1-Δq11, la cual carece del exón 11, se localiza en el citoplasma de células tumorales (Wilson y cols., 1997). Otra posible explicación de la expresión de BRCA1 en el citoplasma, implica a la proteína BARD1, la cual interactúa con BRCA1 a través del dominio RING, y promueve la retención nuclear de BRCA1 a través del enmascaramiento de la señal de 20
44 exporte nuclear (NES) de BRCA1 (Fabbro y cols., 2002). Así, la formación del heterodímero BRCA1/BARD1 favorece la localización nuclear de BRCA1. A este respecto, se ha sugerido que mutaciones en el gen BRCA1, en el dominio RING pueden afectar esta interacción produciendo la expresión citoplasmática de BRCA Funciones de la proteína BRCA1 y su relación con la inestabilidad genómica en los tumores de mama. El gen BRCA1 contiene 22 exones y codifica para una proteína de 1863aa, con una masa molecular de 220KDa (Miki y cols., 1994). El extremo amino terminal de la proteína BRCA1 contiene un dominio RING, una secuencia de exporte nuclear (NES) y dos secuencias de localización nuclear (NLS). Hacia el carboxilo terminal contiene el dominio BRCT, presente en variadas proteínas que responden al daño del DNA. Este dominio une diferentes proteínas fosforiladas (Miki y cols., 1994; Chen y col., 1996; Thakur y cols., 1997). Variadas funciones han sido atribuidas a BRCA1 en la célula normal. Esta proteína participa en la regulación de la transcripción (asociándose a la RNA polimerasa II), está implicada en el remodelamiento de la cromatina, regula puntos de control del ciclo celular en la fase S y en el paso de la fase G2-M, y presenta actividad ubiquitina-ligasa (a través de su dominio RING, e interactuando con BARD1). Además, participa en la reparación del DNA por quiebres de la doble hebra mediante recombinación homóloga. (Moynahan y cols., 1999; Starita y Parvin, 2003; Deng, 2006). Estas funciones nucleares son relevantes en el mantenimiento de la integridad genómica de la célula (Li y Greenberg, 2012). En relación, a la reparación del DNA, se han descrito tres mecanismos diferentes de reparación del DNA por quiebres de la doble hebra. La recombinación no homóloga ( nonhomologous end joining (NHEJ), la recombinación homóloga (HR) y por el apareamiento de la 21
45 hebra simple ( single strand annealing o SSA). De éstos, el mecanismo predilecto durante las fases S y G2 del ciclo celular corresponde a la recombinación homóloga, dependiente de BRCA1, y que es libre de errores en comparación con los otros dos mecanismos. Por lo tanto, cuando la función de BRCA1 está ausente, la célula no puede usar el mecanismo de recombinación homóloga y se utiliza uno de los otros mecanismos alternativos de reparación del DNA (NHEJ y SSA), los cuales son capaces de producir errores en el DNA. De esta manera, se ha sugerido que aumenta la presencia de mutaciones, translocaciones e inestabilidad cromosomal como consecuencia de la pérdida de función de BRCA1 en la célula. (Turner y cols., 2004; Kennedy y cols., 2004). Esto lleva a una inestabilidad genómica que se manifiesta en un aumento de la tasa de alteraciones genómicas, de ganancias y pérdidas de regiones del DNA que pueden contener genes implicados en cáncer como oncogenes o genes supresores de tumor. (Kwei y cols., 2010). Un aspecto relevante de las células deficientes de BRCA1, o de BRCA2 que también participan en la reparación del DNA, es que son especialmente sensibles a apoptosis luego de ser tratadas con inhibidores de las enzimas reparadoras poli [ADP]-ribosa polimerasa 1 y 2 (PARP1 y PARP2), cuando estos inhibidores son agregados luego de un daño en el DNA. Ensayos clínicos en fase I en pacientes que portan mutaciones en los genes BRCA1 o BRCA2 en la línea germinal, utilizando el fármaco Olaparib, inhibidor de PARP1, mostró una fuerte actividad antitumoral (Fong y cols., 2009). PARP1 es una enzima localizada en el núcleo que participa en la reparación del quiebre de la simple hebra del DNA a través de la reparación por escisión de base (Woodhouse y Dianov, 2008; Caldecott, 2007). Cuando PARP1 es inhibida se generan quiebres de la doble hebra de DNA durante la replicación (Woodhouse y cols., 2008), los cuales deberían ser reparados por el sistema de reparación del quiebre de la doble hebra por recombinación homóloga, que involucran a BRCA1 y BRCA2. 22
46 13. Perfiles de rearreglos genómicos, mediante array-cgh, en cáncer de mama. La inestabilidad genómica es una característica de varios tipos de cáncer humanos, incluido el cáncer de mama. Es producto de la acumulación de alteraciones genómicas y fuente de heterogeneidad inter e intra-tumor, generando poblaciones de células tumorales que pueden estar sujetas a selección en un microambiente dado (Burrell y cols., 2013). Está, se puede apreciar en términos de la inestabilidad cromosomal, que produce aneupliodía, es decir, desbalance en el número de copias de los cromosomas. La inestabilidad, puede deberse a una disfunción de los cromosomas, disfunción en la segregación durante la mitosis, disfunción de los telómeros, o por acumulación de mutaciones en genes que participan en la reparación del DNA, como mutaciones en el gen BRCA1, previamente mencionado (Kwei y cols., 2010, Burrell y cols., 2013, Huen y cols., 2010). Las alteraciones cromosómicas estructurales pueden involucrar cromosomas completos, brazos cromosómicos, o regiones intra-cromosoma, implicando la pérdida o ganancia de información génica relevante contenidos en ellos (Burrell y cols., 2013). Se define que la ganancia de una región cromosómica puede conllevar el aumento de la actividad de un protoncogen, en cambio, la pérdida de una región cromosomal, la cual puede contener un gen supresor de tumor, podría dar lugar a la inactivación de éste. Así, la acumulación de alteraciones genómicas puede afectar distintas funciones celulares, que pueden conferir a la célula un fenotipo transformador y finalmente darle la capacidad de progresar en el desarrollo del cáncer. La identificación de regiones cromosómicas con cambios en su número de copias puede facilitar la localización e identificación de oncogenes o genes supresores de tumor, o bien vías de señalización, que pueden corresponder a potenciales blancos de tratamiento en cáncer de mama (Pollack y cols., 2002). 23
47 El primer método molecular que permitió un análisis sistémico de los cambios cromosomales, a través del genoma entero fue la Hibridación Genómica Comparativa (CGH) en cromosomas en metafase. Mediante este método se caracterizó el genoma de varios tipos de cáncer, sin embargo, su limitante es la baja resolución. (Albertson y cols., 2003) Con el desarrollo de tecnologías de mayor resolución, surgen los microarreglos de DNA, que pueden ser hibridados con DNA genómico proveniente de un tejido específico, llamado array- CGH (microarreglos de Hibridación Genómica Comparativa) o con RNA, denominado microarreglos de expresión génica. Para el estudio de biopsias fijadas en formalina y embebidas en parafina, la técnica predilecta es el array-cgh, ya que el DNA es más estable que el RNA en las condiciones en que se encuentra inmerso el tumor (van Beers y Nederlof., 2006). Con esta técnica, los tumores son caracterizados en relación a las alteraciones cromosomales estructurales desbalanceadas en el genoma, como pérdidas de ambos alelos (0 copias), pérdidas de un alelo (1 copia), ganancias (3-5 copias), y amplificaciones ( 5 copias) (van Beers y Nederlof., 2006). El uso de array-cgh, en cáncer de mama, ha permitido identificar grandes alteraciones cromosomales y rearreglos genómicos de menor longitud cuyos genes contenidos pueden contribuir a la formación del tumor maligno (Chao y cols., 2012). Evidencias revelan que alteraciones en el número de copias (rearreglos genómicos) pueden llevar directamente a la desregulación global de la expresión génica, mecanismo, por el cual, las células se convierten en tumorigénicas (Pollack y cols., 2002; Hyman y cols., 2002; Chao y cols., 2013). A este respecto, entre un 20-62% de los genes localizados en regiones genómica amplificadas, están sobreexpresados en cáncer de mama (Hyman y cols., 2002; Pollack y cols., 2002; Turner y cols., 2010). De este modo, se han identificado conocidos oncogenes: FGFR1 (8q11), MYC (8q24), CCND1 (11q13), ERBB2 (17q22), y ZNF217 (20q13), implicados en cáncer de mama. Por otra parte, en 24
48 regiones genómicas que presentan deleción se identificaron, los genes supresores de tumor RB1 (13q14), y TP53 (17p13), cuyas vías de señalización están involucradas en el desarrollo de diferentes grupos de tumores de mama. (Pollack y cols., 2002; Turner y cols., 2010). Por otra parte, basándose en la selección de los tumores de mama por su inmunofenotipo, perfil de expresión molecular o predisposición genética, se han identificado cambios en el número de copias genómicos específicos para un grupo de tumor definido. Esto sugiere, que específicas vías de señalización o procesos biológicos están implicados en el desarrollo y fenotipo observado en un grupo definido de tumores. Así, estudios en tumores de mama esporádicos han permitido identificar diferentes deleciones y amplificaciones genómicas, según el estado del receptor de estrógeno e incluso con el tipo histológico de los tumores. (Albertson, 2003; Reis-Filho y cols., 2005; Bergamaschi y cols, 2006; Chin y cols, 2007). De la misma forma, tumores de mama provenientes de pacientes con cáncer de mama hereditario, revelan que los tumores de pacientes portadoras de una mutación en el gen BRCA1 por la línea germinal, presentan deleciones y amplificaciones génicas comunes, que difieren de los tumores de pacientes que portan mutación en el gen BRCA2 y de los tumores esporádicos (Jonsson y cols, 2005; Melchor y cols, 2005; Álvarez y cols, 2005). A este respecto, se ha descrito que los tumores con mutación en el gen BRCA1 portan rearreglos genómicos específicos de ganancia: 3q27.1-q27.33, 7q36.1-q36.3, 10p15.3, 10p15.1-p14, amplificación de la región 6q que contiene el protoncogen MYB, implicado en la progresión del cliclo celular de G2/M y en la estabilidad genómica. Este protoncogen se ha encontrado amplificado en el 29% de los tumores con mutación en BRCA1 (Kauraniemi y cols., 2000). Además, deleción de las regiones cromosómicas 5q11.2-q23.8, 15q , 15q.3-q21.1 (Jonsson y cols., 2005). En cambio, los tumores de portadoras de mutación en el gen BRCA2, presentan ganancias específicas de las regiones: 17q23.3-q24.2, 20q12-q13.2, y amplificación de 25
49 la región 12q14.2-q21.1 donde se localiza el protoncogen MDM2. A su vez, de deleciones genómicas que corresponden a: 11q14.3-q21 y 11q24.2-q25 (Jonsson y cols., 2005; Álvarez y cols., 2005; Melchor y cols., 2005). 14. Rearreglos genómicos en tumores de mama triple negativo y tumores del grupo tipo Basal. En tumores de cáncer de mama con fenotipo triple negativo, Han y cols., (2008) identificaron un perfil de rearreglos genómicos de ganancias recurrentes entre ellos, que involucra las regiones cromosómicas: 5q15.33, 8q24.3, 9q24.3-p21.1, 10p15.3-p14, 12p13.33-p13.31, 20q13.33 y 21q22.3. Estos rearreglos genómicos fueron poco frecuentes en tumores que expresan los receptores hormonales y en aquellos que presentan amplificación de HER2+ (Han y cols, 2008). Los autores, interceptaron los genes contenidos en las regiones genómicas de ganancia recurrentes en los tumores triple negativo con dos bases de datos de expresión génica de tumores tipo Basal, publicadas (Kapp y cols., 2006 y de Richardson y cols., 2006). Asi, usando esta aproximación indirecta, identificaron potenciales genes que se encuentran altamente expresados en tumores de cáncer de mama esporádicos tipo Basal. Sugiriendo, que su aumento de expresión se correlaciona con la ganancia genómica observada para el gen (Kapp y cols., 2006 y de Richardson y cols., 2006). Este estudio, sugiere que este grupo de tumores de mama acumulan ganancias genómicas diferente, en relación a otros tipos de cáncer de mama, y cuyos genes contenidos pueden ser relevantes para el conocimiento de la biología de estos tumores. Otro estudio en tumores de mama triple negativo, realizado por Andre y cols., (2009), describe nuevas alteraciones genómicas recurrentes en este grupo de tumores, que corresponden a ganancias en la región: 6p21 y 6p23, y las regiones genómicas de deleción: 15q14 y 15q22. Aún 26
50 más, mediante un agrupamiento jerárquico no supervisado de los datos de array-cgh, provenientes de los tumores triple negativo y de otros grupos de tumores de mama, encontraron que el rearreglo genómico 6p21.2-p12 permite distinguir a un 64% de sus tumores triple negativo en comparación con el resto de los tumores de mama (Andre y cols., 2009). Por su parte, Turner y cols., (2010) mediante el análisis por array-cgh y microarreglos de expresión génica en tumores de mama triple negativo, identificó genes sobre-expresados cuando éstos se encontraban amplificados. Estas regiones corresponden a 10q26.3 (FGFR2 y BUB3), 13q34 (RAB20), 19p13.12 (PKN1 y NOTCH3). Por otra parte, los estudios de array-cgh en tumores de mama tipo Basal comparado con tumores de mama de otros subtipos han identificado rearreglos genómicos específicos para el subgrupo tipo Basal. Uno de estos rearreglos, consiste en la ganancia de la región 10p, la cual contiene los genes MAP3K8, ZEB1 y FAM107B (Adelaide y cols., 2007; Weigman y cols., 2012), además de la pérdida de la región 16p (Haverty y cols., 2009), deleción de la región 5q11-35, que contiene los genes RAD17 y RAD50, implicados en la reparación del DNA, dependiente de BRCA1 (Bergamaschi y cols., 2006; Weigman y cols., 2012). Por último, deleción de la región 4q que contiene el gen INPP4B, un gen supresor de tumor, implicado en la inhibición de la señalización de PI3K (Weigman y cols., 2012). 27
51 Hipótesis Tumores de cáncer de mama con fenotipo triple negativo/tipo basal y pérdida de expresión de la proteína BRCA1, comparten un patrón similar de ganancias y pérdidas genómicas. Objetivo general Analizar tumores de cáncer de mama con fenotipo triple negativo/ tipo Basal y con pérdida expresión de la proteína BRCA1 mediante array-cgh, con el fin de identificar un patrón de regiones de ganancias y pérdidas genómicas que pueden contener potenciales genes supresores de tumor y protoncogenes, involucrados en el desarrollo del cáncer y que en un futuro pueden constituir blancos de terapia. Objetivos específicos 1. Caracterizar los tumores mamarios con fenotipo triple negativo en relación al fenotipo tipo Basal 2. Evaluar la expresión de la proteína BRCA1 en tumores con fenotipo triple negativo/ tipo Basal 3. Identificar regiones de ganancias y pérdidas genómicas recurrentes en los tumores de cáncer de mama triple negativo/ tipo Basal con pérdida de expresión de BRCA1. 28
52 4. Comparar el patrón genómico identificado en los tumores de cáncer de mama triple negativo/ tipo Basal con pérdida de expresión de BRCA1 con los datos publicados en relación a otros tipos de tumores de mama. 5. Identificar en los tumores de cáncer de mama triple negativo/ tipo Basal con pérdida de expresión de BRCA1, protoncogenes y genes supresores de tumor localizados en las regiones de ganancias y pérdidas genómicas recurrentes entre ellos, y además determinar los posibles procesos biológicos alterados. 29
53 1. PACIENTES 1.1. Pacientes del estudio Las pacientes ingresadas a este estudio presentaron cáncer de mama y desarrollaron un tipo particular de tumor, denominado triple negativo. Este fenotipo de tumor de mama se caracteriza por la ausencia de expresión del receptor de estrógeno (RE), del receptor de progesterona (RP) y del receptor 2 del factor de crecimiento epidermal (HER2). Estas pacientes fueron derivadas desde cinco centros hospitalarios, localizados en la ciudad de Santiago, Región Metropolitana de Chile. Estos son: el hospital Clínico de la Pontificia Universidad Católica de Chile (PUC), el hospital Barros Luco (HBL), el hospital San Borja Arriarán (HSBA), el hospital José Joaquín Aguirre (JJA) y el Instituto Nacional del Cáncer (INC). Cabe hacer notar, que las pacientes proceden mayoritariamente de los primeros dos centros hospitalarios. Las pacientes y esta investigación doctoral, están incluidas dentro del proyecto de investigación, financiado por FONDECYT (# ) a cargo de la Dra. Pilar Carvallo. Para desarrollar esta investigación seleccionamos 46 pacientes con cáncer de mama, prevenientes de 43 familias no relacionadas. A partir de ellas, obtuvimos una muestra de su tumor de mama fijada en formalina y embebidos en parafina, y una muestra de sangre periférica para extraer el DNA genómico desde los linfocitos. Esto último, corresponde a un objetivo del proyecto FONDECYT, cuya finalidad es buscar mutaciones en los genes BRCA1 y BRCA2 en la línea germinal. El protocolo del consentimiento informado se encuentra aprobado por el Comité de Ética de la Facultad de Medicina de la Pontificia Universidad Católica de Chile y por el Comité Ético-Científico del Servicio de Salud Metropolitano Central. 30
54 1.2. Selección de las pacientes con tumores de mama triple negativo. Las pacientes con cáncer de mama cuyo examen histopatológico del tumor, realizado por un médico anátomo-patólogo del hospital respectivo, reveló ausencia de expresión del receptor de estrógeno (RE-), del receptor de progesterona (RP-) y no expresión del receptor 2 del factor de crecimiento epidermal (HER2), son designados con fenotipo triple negativo. Estos tumores son llamados, generalmente en la literatura, como TNBC, sigla que deriva de la frase en inglés: Triple Negative Breast Cancer, y que utilizaremos para referirnos a estos tumores en el escrito. A partir de estos antecedentes, solicitamos tumores fijados en formalina y embebidos en parafina (FFEP), con sus respectivos informes histopatológicos a cada unidad de anatomía patológica del hospital correspondiente. A partir de las 46 pacientes obtuvimos 48 tumores TNBC, ya que, una paciente presentó dos tumores uno en cada mama (bilateral) y otra paciente desarrolló dos tumores primarios, en la misma mama, identificados por el anátomo-patólogo correspondiente. En ambos casos, los tumores fueron usados como muestras independientes. A continuación, en la tabla 1, se muestran las principales características clínicas e histopatológicas de los tumores TNBC. En esta tabla, está registrada la edad a la cual la paciente es diagnosticada con el cáncer de mama y el hospital del cual provienen, la histología del tumor y el tipo histológico, y finalmente el grado del tumor (sólo cuando se encuentra indicado en el informe de biopsia de la paciente entregado por el centro hospitalario del cual deriva). Además, se indican aquellas pacientes que son portadoras de una mutación en el gen BRCA1 o BRCA2 en la línea germinal, identificadas según el objetivo del proyecto FONDECYT. 31
55 Tabla 1: Características clínico-patológicas de las pacientes chilenas y de sus tumores TNBC, ingresadas al estudio. N Hospital ID Familia ID Tumor Edad de diagnóstico Mutación BRCA1 BRCA2 Histología del tumor Tipo histológico del tumor Grado del tumor 1 PUC TN01 T_TN01 26 Carcinoma ductal invasor Medular ND 2 PUC TN07 T_TN07 30 Carcinoma ductal invasor Medular ND 3 PUC TN11 T_TN11 33 Carcinoma ductal invasor Medular III 4 PUC TN16 T_TN16 30 Carcinoma ductal invasor Medular ND 5 PUC TN17 T_TN17 30 Si Carcinoma ductal invasor Mucinoso III 6 PUC TN12 T_TN12 32 Carcinoma ductal invasor Metaplásico II 7 PUC T_TN21_1 50 Carcinoma ductal invasor Metaplásico III TN21 PUC T_TN21_2 50 Carcinoma ductal invasor Metaplásico III 8 PUC TN08 T_TN08 49 Carcinoma ductal invasor Papilar ND 9 PUC TN15 T_TN15 60 Carcinoma ductal invasor Tubular I 10 PUC TN03 T_TN03 33 Carcinoma ductal invasor no especial II 11 PUC TN19 T_TN19 62 Carcinoma ductal invasor no especial II 12 PUC TN20 T_TN20 40 Carcinoma ductal invasor no especial II 13 PUC TN46 T_TN46 65 Carcinoma ductal invasor no especial II 14 JJA F61 T24 33 Si Carcinoma ductal invasor no especial II PUC T41 33 Carcinoma ductal invasor no especial III 15 PUC F77 T42 31 Carcinoma ductal invasor no especial III PUC T43 58 Carcinoma ductal invasor no especial III 16 PUC TN02 T_TN02 29 Carcinoma ductal invasor no especial III 17 INC F24 T5 28 Si Carcinoma ductal invasor no especial III 18 PUC FP4 T_P4 22 Si Carcinoma ductal invasor no especial III 19 PUC F78 T_M Si Carcinoma ductal invasor no especial III 20 PUC F76 T39 43 Carcinoma ductal invasor no especial III 21 INC F42 T11 62 Carcinoma ductal invasor no especial III 22 PUC TN10 T_TN10 58 Si Carcinoma ductal invasor no especial III 23 PUC T_TN24 38 Carcinoma ductal invasor no especial III TN24 PUC T_TN44 41 Carcinoma ductal invasor no especial III 24 HSBA TN39 T_TN39 62 Si Carcinoma ductal invasor no especial III 25 PUC TN14 T_TN14 65 Carcinoma ductal invasor no especial III 26 PUC TN34 T_TN34 47 Carcinoma ductal invasor no especial III 27 HSBA TN38 T_TN38 45 Carcinoma ductal invasor no especial III 28 HBL TN31 T_TN31 45 Carcinoma ductal invasor no especial III 29 PUC TN43 T_TN43 77 Carcinoma ductal invasor no especial III 30 HBL TN49 T_TN49 76 Carcinoma ductal invasor no especial III 31 HBL TN27 T_TN27 39 Carcinoma ductal invasor no especial III 32 PUC TN04 T_TN04 55 Carcinoma ductal invasor no especial III 33 HBL TN32 T_TN32 50 Carcinoma ductal invasor no especial III 34 PUC TN45 T_TN45 53 Carcinoma ductal invasor no especial III 35 PUC TN47 T_TN47 66 Carcinoma ductal invasor no especial III 36 PUC TN48 T_TN48 61 Carcinoma ductal invasor no especial III 37 HBL TN50 T_TN50 67 Carcinoma ductal invasor no especial III 38 HBL TN51 T_TN51 42 Carcinoma ductal invasor no especial III 39 PUC T_TN06_1 42 Carcinoma ductal invasor no especial III TN06 PUC T_TN06_2 54 Carcinoma ductal in situ NA NA 40 HBL TN28 T_TN28 58 Carcinoma ductal in situ NA NA 42 PUC TN05 T_TN05 41 Carcinoma ductal in situ NA NA 42 PUC TN42 T_TN42 55 Carcinoma ductal in situ NA NA 43 PUC TN09 T_TN09 45 Carcinoma lobulillar invasor NA NA En esta tabla, se muestra el hospital de donde provienen las pacientes, tal como se indica en el texto, los identificadores de la familia y del tumor y la edad de diagnóstico del cáncer de mama. Además, se indica la histología del tumor, y el tipo histológico, el grado histológico del tumor y finalmente la presencia de una mutación en los genes BRCA1 o BRCA2 en la línea germinal. Subíndice a: indica la paciente con cáncer de mama bilateral, b: paciente con dos tumores primarios, c: familias con más de un familiar con cáncer de mama con fenotipo triple negativo. NA: No aplica, nd: No definido en el informe histopatológico de la biopsia realizada a la paciente en cuestión. 32
56 1.3. Criterios para seleccionar familias con cáncer de mama hereditario. Las pacientes seleccionadas por el fenotipo del tumor son encuestadas, con el fin de determinar si tienen historia familiar de cáncer de mama y/u ovario hereditario, ya sea por la línea paterna o la línea materna, o bien si presentan otros factores hereditarios como la edad muy temprana de aparición del cáncer (menor de 40 años), la bilateralidad y la presencia de cáncer de mama y de ovario en la misma paciente. El cáncer se define como hereditario, si la familia cumple, al menos uno de los siguientes criterios: 1. tres familiares con cáncer de mama a cualquier edad, en una o más generaciones. 2. dos familiares con cáncer de mama, una de ellas antes de los 43 años (premenopáusico) en una o más generaciones. 3. dos familiares afectados de cáncer, una de mama y otra de ovario a cualquier edad, en una o más generaciones. 4. dos familiares con cáncer de mama uno de ellos hombre, en una o más generaciones. En relación a estos criterios, sumado a las pacientes seleccionadas por presentan factores hereditarios previamente mencionados, encontramos que un 67% (29/43) de las pacientes seleccionadas por tumores triple negativo correspondía a uno de los criterios de cáncer hereditario y un 33% (14/43) no cumplían. 33
57 2. MATERIALES 2.1. Material químico, enzimas y anticuerpos Los reactivos sólidos y líquidos, enzimas y anticuerpos utilizados para el desarrollo de esta investigación se obtuvieron de las siguientes empresas proveedoras. De Merck, se obtuvo el NaCl, el etanol 100%, el ácido bórico, el Neo Clear (sustituto del xileno), el PBS 10X, el citrato de sodio, la hematoxilina Mayers y el medio de montaje acuoso. De Bio-Rad, se obtuvo el Tween-20 y las columnas cromatográficas para purificación de DNA Micro-BioSpin. De Fermentas, se obtuvieron las enzimas de restricción AluI y RsaI, necesarias para la digestión enzimática del DNA genómico tumoral amplificado con la DNA polimerasa del fago phi29. De Promega, se obtuvo la proteinasa K, utilizada para extraer el DNA genómico proveniente de las células tumorales microdisectadas. De Agilent Technologies, se obtuvieron las plataformas en que se realizó la técnica de array- CGH. Estas plataformas de microarreglos de array-cgh corresponden al Human Genome CGH 4x44K. Además, obtuvimos el sistema de hibridación oligo acgh/chip-on-chip hybrization kit, la solución estabilizadora, y el acetonitrilo. De GE Healthcare, se obtuvieron los fluoróforos Cy3dUTP y el Cy5dUTP, y el sistema Genomiphi HY DNA Amplification kit. Este último, contiene la DNA polimeras del fago phi29 y los partidores al azar, que amplifican el DNA genómico tumoral. De Invitrogen, se obtuvo el EDTA, el Tris-HCl, el N-lauroilsarcosina, el DNA repetitivo humano Cot-1, el sistema de marcación Bioprime arraycgh labeling Module, usados en la técnica de array-cgh. Además, de esta empresa obtuvimos el anticuerpo monoclonal anti-egfr (clon 34
58 31G7), la enzima Pepsina, y el sistema de tinción Histostain-Plus, usados para desarrollar la técnica de inmunohistoquímica. De Sigma Chemicals Co, se obtuvo el fosfato de sodio monobásico. De Chemicon, se obtuvo el anticuerpo monoclonal anti-brca1 (clon MS110). De Novocastra, se obtuvo el anticuerpo monoclonal anti-ck14 (clon LL002). De Thermo scientific, se obtuvo el anticuerpo monoclonal anti-ck5 (clon XM26) De Life Technologies, se obtuvieron los reactivos e insumos necesarios para realizar la microdisección por captura con láser de células tumorales. Estos reactivos son: la solución de tinción de las células tumorales, denominada Histogene, el amortiguador de extracción de las células desde el polímero. Además, de los polímeros termolábiles, llamados macrocaps, donde se adhieren la células tumorales al atravesar el láser el tejido. 35
59 3. METODOS Análisis de la expresión de las citoqueratinas 5, 14, y del receptor 1 de EGF y de la proteína BRCA1 en tumores TNBC mediante inmunohistoquímica. La detección de las citoqueratinas 5 (CK5), 14 (CK14), y del receptor 1 del factor de crecimiento epidermal (EGFR) y de la proteína BRCA1 son realizadas por la técnica de inmunohistoquímica en secciones de tejido fijado en formalina y embebido en parafina (FFPE). Para esto, se utilizan cortes histológicos seriados de 5µm de espesor montados en portaobjetos (con xilano) de cada tumor de mama (n=48). Además, se utilizan cortes histológicos de igual espesor de un microarreglo de tejido construido con 24 de los 48 tumores de mama de este estudio. Brevemente, el microarreglo es diseñado en el Departamento de Anatomía Patológica de la Pontificia Universidad Católica de Chile. Para esto, se extraen cilindros de 0,2 mm de diámetro desde regiones representativas del tumor de cada una de las muestras y se organizan en un recipiente para ser embebidos en parafina. En este microarreglo, se utilizaron como controles positivos tejidos de mama normal, piel y amígdala. Los tres tejidos de mama normal provienen de reducciones mamarias realizadas a mujeres sin cáncer de mama. Estos son utilizados para analizar la expresión normal de las citoqueratinas y de BRCA1. Por otra parte, el tejido de amígdala se usó como control positivo de la expresión de la CK5. Para la detección de la CK14 se utilizó el tejido procedente de piel. El control negativo corresponde a un tejido sin incubación con el anticuerpo primario. La tinción de los cortes se realiza usando el sistema comercial Histostain-Plus. Inicialmente, los cortes histológicos montados en los portaobjetos son desparafinados, incubándolos a 70ºC por 30 minutos y lavados en tres soluciones de Neo-Clear (un derivado del xileno, no tóxico) cada 5 minutos. La re-hidratación se realiza incubando los portaobjetos dos veces en soluciones de etanol al 100, 95, y al 70% respectivamente, durante 5 minutos. A 36
60 continuación, los cortes histológicos son lavados en PBS 1X durante 10 minutos. Luego, se inactiva la peroxidasa endógena con una solución de H202 al 3% en etanol al 70% durante 30 minutos. Posteriormente, los tejidos son lavados tres veces en PBS 1X por 5 minutos. La exposición del antígeno, para las citoqueratinas y la proteína BRCA1 se realiza, incubándolos en una solución de Citrato Sodio 10 mm, ph 6,0 y Tween 20 al 0,1 %, en un horno microonda por 20 minutos (4 minutos con potencia al 100% y 16 minutos con potencia al 20%). Posteriormente, estos se enfrían a temperatura ambiente durante 20 minutos e inmediatamente son lavados tres veces en PBS 1X, por 5 minutos. Por otra parte, la exposición del antígeno de EGFR, se realiza mediante digestión enzimática con la solución de Pepsina. Para esto, los portaobjetos se colocan en una cámara húmeda y se incuban a 37 C durante 20 minutos en un horno de hibridación. A continuación, de la exposición del antígeno, los portaobjetos se colocan en una cámara húmeda y son incubados con la solución bloqueadora de suero de cabra al 10% (Histostain-plus) durante 60 minutos. Después, son incubados con el anticuerpo primario correspondiente. Para detectar la CK5 se utiliza el anticuerpo monoclonal anti-ck5, clon XM26 a una dilución de 1:50. Para la CK14 se usa el anticuerpo monoclonal anti-ck14, clon LL002 a una dilución de 1:100. Por su parte, para EGFR se utiliza el anticuerpo monoclonal anti-egfr, clon 31G7 a una dilución de 1:50. Finalmente, para la detección de BRCA1 se utiliza el anticuerpo monoclonal anti-brca1 a una dilución de 1:50. Este anticuerpo reconoce un epítope de los primeros 300 aminoácidos, ubicados en el N-terminal de la proteína. A continuación, los cortes son colocados en la cámara húmeda y se dejan a 4 C durante la noche. Al día siguiente, los cortes son lavados tres veces en PBS 1X, durante 5 minutos e incubados con el anticuerpo secundario biotinilado universal (Histostain-plus) durante 60 minutos. Luego, son lavados tres veces en PBS 1X por 5 minutos y se incuban durante 30 minutos con el reactivo conjugado compuesto de la enzima peroxidasa- estreptavidina y se 37
61 lavan con PBS 1X tres veces por 5 minutos. Finalmente, las muestras se exponen al cromógeno 3- amino-9-etilcarbazol (AEC) durante 15 minutos en una cámara húmeda, el cual produce un precipitado color rojo. Excepción a esto, son 30 muestras incubadas con el anticuerpo anti-ck5 las cuales son expuestas al cromógeno 3,3-Diaminobenzidina (DAB), el que produce un precipitado color café. Para el contraste nuclear se utiliza la tinción con hematoxilina de Mayer. Esta tinción permite, teñir los núcleos de color celeste. Para esto, los porta-objetos se incuban 5 segundos con el reactivo y son incubados en agua potable durante 1 minuto, luego son lavados con PBS 1X. Finalmente, se adiciona una gota de medio de montaje acuoso y son cubiertos con un cubre-objeto. De esta manera, las secciones de tejidos teñidas son observadas en un microscopio de luz, Olympus CX31. Las imágenes son visualizadas con el software Micrometrics TM SE Premium, versión 2.8 y obtenidas con la cámara Olympus, modelo CD Interpretación de la detección de las citoqueratinas, de EGFR y de la proteína BRCA1 en los tumores TNBC. Con el fin de identificar los tumores positivos para la CK5, la CK14, EGFR y la proteína BRCA1, dos médico-patólogas de la unidad de anatomía patológica del Hospital San Borja Arriarán, en forma independiente y junto a la investigadora de este estudio, realizaron observaciones de las tinciones usando un microscopio de luz. Estas observaciones comprenden la estimación de la intensidad de la tinción, el cálculo del porcentaje de células tumorales, y la localización subcelular de la marca. Además, se determina la distribución de la tinción para cada marcador, en el tejido tumoral (heterogéneo/homogéneo). 38
62 a. Tumores positivos para las CK5 y CK14. En estos dos casos, los tumores TNBC son positivos si se observa inmunoreactividad de los anticuerpos anti-ck5 y anti-ck14 en igual o mayor al 1% de las células neoplásicas. Para esto, se contabilizan las células tumorales en las regiones del corte histológico con mayor densidad de células neoplásicas. La intensidad de tinción en estos tumores es estimada, basándose en la intensidad del color del cromógeno como ausente, débil, moderado o fuerte. De esta manera los tumores son agrupados en cuatro categorías: 1) 0, corresponde a tumores sin detección del anticuerpo y denominados negativos para la expresión del marcador proteico correspondiente. 2) 1+ tumores positivos, con intensidad de tinción débil, de la proteína respectiva. 3) 2+ tumores positivos con una moderada intensidad de tinción y por último 4) 3+ tumores positivos con fuerte intensidad de tinción. Además, se observa que la localización de la tinción de estas citoqueratinas se encuentre en el citoplasma/membrana. Finalmente, determinamos la distribución de la tinción en los tumores TNBC positivos para cualquiera de estas citoqueratinas como: Homogénea, si se observa la marca en toda la extensión de la región tumoral, involucrando más del 90% de las células neoplásicas con expresión del marcador proteico. Heterogéneo, si se observan regiones del tumor con células positivas y células negativas en la región de alta densidad de células neoplásicas. Además, se determina si es focalizada, comprendiendo regiones tumorales con zonas positivas y zonas con células neoplásicas negativas, o bien dispersas, donde se observa que en toda la extensión del tumor, existen células tumorales positivas mezcladas con células tumorales negativas. b. Tumores positivos para EGFR. Se ha descrito que los tumores TNBC son positivos para EGFR, en más del 10% de las células tumorales, si se observa inmunoreactividad con el anticuerpo. Tal como está descrito para 39
63 las citoqueratinas, se estimó la intensidad de tinción como ausente (0), débil (1+), moderado (2+) o fuerte (3+) en cada caso. En relación a la localización de EGFR se determinó si la marca se encuentra en el citoplasma y/o membrana, en cada tumor. Además, se determinó la distribución de la tinción heterogénea u homogénea, igual que para las citoqueratinas. c. Tumores positivos para BRCA1. En este caso, definimos que los tumores son positivos para BRCA1 si en igual o mayor al 10% de las células tumorales se observa tinción. En estos tumores es muy importante determinar la intensidad de la tinción y la localización de la marca. De esta manera, igual a los casos anteriores, se estiman los tumores con intensidad de tinción débil (1+), aquellos con intensidad de tinción moderada (2+) y los que tienen intensidad de tinción fuerte (3+), además, de los negativos en el área tumoral (0). En relación a la localización de la marca, se determina si los tumores están expresando en el núcleo y/o citoplasma de las células tumorales. Además, se observa si la distribución de la tinción es heterogénea u homogénea Valorización de la tinción, calculando el puntaje-h. Utilizando la estimación de la intensidad de tinción y el porcentaje de células tumorales con inmunoreactividad para cada marcador se realiza una valorización de la tinción, determinando el puntaje-h, el cual es calculado como el producto entre la intensidad de la tinción y el porcentaje de las células positivas. La fórmula del puntaje-h es la siguiente: (% de células negativas) x 0 + (% de células con intensidad 1+) x 1 + (% de células con intensidad 2+) x 2 + (% de células con intensidad 3+) x 3. Así, este puntaje-h proporciona una variable continua cuyo rango es de 0 a 300 y que permite realizar comparaciones entre los diferentes marcadores proteicos evaluados en este estudio. 40
64 3.1.3 Definición de los tumores TNBC, en tipo basal y no tipo basal Con la finalidad de determinar si los tumores TNBC corresponden al tipo Basal o al tipo no-basal, se utiliza la expresión de las citoqueratinas y de EGFR. Este panel de marcadores proteicos, mediante inmunohistoquímica, es usado y descrito ampliamente en la literatura, como se mencionó en la introducción. Para esto, los tumores TNBC/tipo Basal se definen si expresan al menos la CK5 y/o la CK14 y/o EGFR. En cambio, los tumores TNBC/tipo no-basal son aquellos que son negativos para la CK5, la CK14 y EGFR Preparación del DNA genómico desde células tumorales. A partir del tumor de mama, fijado en formalina y embebido en parafina, de cada paciente seleccionada se microdisectan las células neoplásicas y se extrae el DNA genómico tumoral Identificación de las células tumorales y microdisección Con el fin de extraer el DNA genómico desde las células tumorales de los tejidos de mama, el médico patólogo, con un microscopio de luz, identifica el área tumoral en cada uno de los cortes histológicos teñidos con hematoxilina y eosina. El requisito es que el área seleccionada sea densa en células tumorales, con más de un 80% de células neoplásicas. Luego, el corte teñido con el área tumoral demarcada, es utilizado como referencia para identificar el área tumoral en los cortes seriados adicionales, teñidos con la solución de tinción histogene. Las células tumorales fueron obtenidas mediante dos métodos de microdisección, uno manual, usado en los primeros 17 tumores TNBC y la microdisección con captura por láser, realizado en los 31 casos restantes. La finalidad de microdisectar células tumorales es enriquecer 41
65 la muestra con el DNA genómico tumoral y no obtener mezcla con otros tipos celulares que pueden estar presentes en el corte histológico, como por ejemplo linfocitos y fibroblastos. a. Microdisección manual de las células tumorales Los cortes histológicos seriados de 5 µm de espesor son teñidos con hematoxilina y procesados por una tecnóloga médica del Laboratorio de Anatomía Patológica de la Pontificia Universidad Católica de Chile. Mediante este método se extrajeron entre a células tumorales respectivamente en cada corte histológico. b. Microdisección por captura con láser de células tumorales. Esta técnica permite aislar poblaciones de células tumorales desde secciones de tejidos heterogéneos. El microdisectador por captura con láser utilizado es el Arcturus (Life thecnology) del Instituto de Investigaciones Materno-Infantil (IDIMI), del hospital del San Borja Arriarán de la Universidad de Chile. Brevemente, se realizaron cortes histológicos seriados de 5 µm de espesor, los cuales son teñidos con la solución de histogene, luego los cortes histológicos son colocados en un desecador durante 30 minutos. Las células tumorales fueron capturadas usando un polímero termolábil, denominado macrocaps. En promedio se obtuvieron a células tumorales por cada tumor por este método Extracción del DNA genómico desde células tumorales microdisectadas manualmente. A partir de las células tumorales, se extrae el DNA genómico tumoral mediante el tratamiento con Proteinasa K y posterior precipitación etanólica. Las células tumorales son incubadas con 300 µl de la solución de digestión con Proteinasa K (Proteinasa K 0,4 µg/µl, EDTA 10mM ph 8, Tris-HCl 20mM ph 8, Tween 20 al 0,4%) por 48 horas a 37 C en un baño con agitación suave. Transcurrido este tiempo, la Proteinasa K se inactiva incubando la mezcla a 95 C 42
66 durante 12 minutos. Luego, las muestras digeridas son centrifugadas para separar el precipitado celular del sobrenadante que contiene el DNA genómico tumoral. El DNA se precipita con 1/10 de volumen de acetato de sodio 3M, ph 5,2 y con 2,5 volúmenes de etanol al 100%. Esta mezcla se incuba durante doce horas a -20 C, y centrifugadas por 40 minutos a velocidad máxima (16.060xg). Por último, el DNA genómico se resuspende en 10 µl de una solución amortiguadora TE-PCR (Tris-HCl 10mM ph 8,0, EDTA 0,1mM). Finalmente, se mide la concentración de DNA en el espectrofotómetro Picodrop Extracción de DNA genómico desde células tumorales microdisectadas por captura con láser. Las células tumorales adheridas en el polímero termolábil (macrocaps), se incuban con el amortiguador de extracción, suministrado por el sistema comercial. Brevemente, 50 µl del amortiguador de extracción se adicionan a un tubo de 0,6 ml y se posicionó el polímero en la boca del tubo. Luego, el tubo se invierte dejando en contacto el líquido con el polímero termolábil. A continuación, se incuban a 42 C en un horno de hibridación durante 30 minutos y se centrifugan a 1.000xg por 5 minutos. El polímero se retira de la boca del tubo y se adicionan 150 µl de la solución de proteinasa K (Proteinasa K 0,4 µg/µl, EDTA 10mM ph 8, Tris-HCl 20mM ph 8, Tween 20 al 0,4%) hasta completar un volumen final de 200 l. La mezcla se incuba durante 48 horas a 37 C en un baño con agitación suave. Posteriormente, se inactiva la Proteinasa K a 95 C durante 12 minutos y se centrifugan a xg por 40 minutos. El DNA se precipita con 1/10 de volumen de acetato de sodio 3M, ph 5,5 y con 2,5 volúmenes de etanol al 100%. Se incuban durante doce horas a -20 C, y luego se centrifugan durante 40 minutos a velocidad máxima (16.060xg). Por último, el DNA se resuspende con un volumen de 10 µl de una solución 43
67 amortiguadora TE-PCR (Tris-HCl 10mM ph 8,0, EDTA 0,1mM). Al final, se cuantifica la concentración del DNA genómico tumoral con el espectrofotómetro Picodrop Hibridación genómica comparativa en microarreglos de DNA (array-cgh) Con el fin de identificar las alteraciones genómicas presentes en los 48 tumores en estudio, se realiza la técnica de hibridación genómica comparativa en microarreglos de DNA (array-cgh). El DNA genómico tumoral se analiza en las plataformas de microarreglos de DNA, Human Genome CGH microarray 4x44K (design ID ), de la empresa Agilent Technologies, Inc. Estos microarreglos contienen sondas correspondientes a oligonucleótidos de 60-mer de longitud, distribuidos a lo largo del genoma completo, de manera que hibridan secuencias codificantes y no codificantes. El promedio de la resolución espacial de cada sonda es de aproximadamente 43Kb (24,2 Kb espaciamiento promedio de las sondas en los genes). Esta plataforma está basada sobre la secuencia de referencia, Human Genome NCBI Build 36 (UCSC hg18, Mar 2006 build). Antes de la hibridación se realiza una mezcla equitativa entre el DNA genómico tumoral del paciente y un DNA genómico de referencia. Cada uno de estos es marcado con fluoróforos diferentes para su posterior identificación. Esta mezcla se hibrida en una plataforma de array-cgh que contiene las sondas que reconocen el genoma completo. El DNA de referencia utilizado corresponde a un pool de DNA genómico extraído desde linfocitos proveniente de 10 hombres sanos al momento de la toma de muestra, obtenidos desde el Banco de Sangre del Hospital Clínico la Pontificia Universidad Católica de Chile. En breve se detallan los distintos pasos involucrados en el análisis mediante la técnica de array-cgh (figura 4) 44
68 3.3.1 Amplificación del DNA genómico tumoral. Con el fin de obtener suficiente cantidad de DNA genómico para el experimento de array- CGH, se realiza la amplificación del DNA genómico tumoral y del DNA genómico de referencia de acuerdo al protocolo descrito por el sistema comercial Genomiphi HY DNA Amplification que utiliza la enzima DNA Polimerasa del fago Phi29. A partir de cada DNA genómico tumoral proveniente de las pacientes con cáncer de mama y de la mezcla de DNA genómico de referencia se realizan diluciones de 8 ng/µl. La amplificación se realiza a partir de 20 ng de DNA genómico tumoral, o del DNA genómico de referencia, contenidos en un volumen final de 2,5 µl respectivamente. Luego, se adicionan 22,5 µl del reactivo amortiguador de la muestra, suministrado por el sistema comercial. A continuación, el DNA genómico se desnatura a 95 C durante 3 minutos e inmediatamente se coloca en hielo. Posteriormente, se adiciona un volumen de 25 µl de una mezcla que contiene el amortiguador de la reacción y la enzima DNA Polimerasa del fago Phi29. La mezcla se incuba a 30 C durante 9 horas. Por último, el proceso se finaliza incubando la mezcla a 65 C durante 10 minutos. La concentración del DNA genómico amplificado es determinada usando un espectrofotómetro Picodrop y la calidad del DNA genómico amplificado se observa en un gel de agarosa al 0,8%, en TBE 1X (Tris-borato 9mM, ph 8,0; EDTA 2mM). 45

69 Figura 4: Representación esquemática de la técnica de Hibridación Genómica Comparativa en microarreglos de DNA o array-cgh. Esta metodología permite identificar deleciones y ganancias genómicas presentes en la muestra de DNA del tumor de mama de la paciente en comparación con una muestra de referencia. Cy3: Cyanine 3 dutp; Cy5: Cyanine 5 dutp. 46
70 3.3.2 Digestión enzimática del DNA genómico Con el objetivo de fragmentar el DNA genómico tumoral y el DNA genómico de referencia se procede a realizar una digestión enzimática. Ésta se realiza con dos enzimas de restricción RsaI y AluI, las cuales fragmentan el DNA en trozos de aproximadamente 500 pb. La digestión se realiza en un volumen final de 100 µl conteniendo 20 µg de DNA, 40U de cada enzima y el amortiguador Tango 1X (Tris-acetato 33mM ph 7,9; acetato de magnesio 10mM; acetato de potasio 66mM; BSA 0,1 mg/ml). El tiempo de incubación de la digestión es de 3 horas a 37 C en un baño termorregulado. Posteriormente, El DNA se precipita con 1/10 de volumen de acetato de sodio 3M, ph 5,2 y con 2,5 volúmenes de etanol al 100%, e incubados durante 12 horas a -20 C. Luego, se centrifugan por 40 minutos a velocidad máxima (16.060xg). El DNA se resuspende con un volumen de 30 µl de una solución amortiguadora TE-PCR (Tris-HCl 10mM ph 8,0, EDTA 0,1mM). El DNA se cuantifica en un espectrofotómetro Picodrop y se observa en un gel de agarosa al 1% en TBE 1X (Tris-borato 9mM, ph 8,0; EDTA 2mM) Marcación del DNA genómico digerido A continuación de la fragmentación del DNA genómico se prosigue con la respectiva marcación. El DNA genómico tumoral se marca con el fluoróforo Cy3 dutp (Cyanine 3 dutp), y el DNA de referencia es marcado con el fluoróforo Cy5 dutp (Cyanine 5 dutp), utilizando el sistema comercial BioPrime array-cgh Genomic Labeling Module. Este sistema se basa en la enzima Klenow (exo -) y partidores al azar para incorporar en la secuencia del DNA sintetizado los nucleótidos marcados con los fluoróforos. En un volumen de 41 µl se realiza la mezcla que contiene 5 µg de DNA y los partidores al azar, la cual es incubada durante 5 minutos a 95 C y luego 5 minutos en frío para su desnaturación. Al DNA desnaturado se le agrega un volumen de 9 47
71 µl de una mezcla que contiene dntps 1X, dutp Cy3 ó Cy5 2pmoles y 40U de enzima Klenow. Esta mezcla se incuba por 9 horas a 37 C en un termociclador. Luego, a la reacción se adicionan 5 µl del amortiguador de EDTA (0,5mM de EDTA ph 8.). Después de la marcación, el DNA es limpiado utilizando columnas Micro-BioSpin (BioRad) de acuerdo a las recomendaciones del proveedor. El DNA marcado se cuantifica en el espectrofotómetro Picodrop, y se calcula la cantidad de DNA efectivamente marcado con cada fluoróforo Cy3 y Cy5.Para esto cada DNA marcado se cuantifica a las longitudes de onda 550nm y 650nm, respectivamente. Finalmente, cantidades equivalentes de DNA marcado son mezclados para la hibridación Hibridación de las muestras Con el fin de identificar las alteraciones genómicas presentes en el DNA de los tumores de mama con fenotipo triple negativo de cada uno de los pacientes, se mezclan cantidades equivalentes de DNA tumoral marcado y del DNA de referencia marcado, las cuales son hibridadas en las plataformas de los microarreglos de DNA Human Genome CGH 4x44. Cada mezcla de DNA se prepara en un volumen final de 110 µl, que contiene 50 g de DNA repetitivo Cot-1 Humano, agente bloqueante 1X (Agilent) y solución amortiguadora de hibridación 1X. Cada mezcla se incuba a 95 C por tres minutos, y luego a 37 C por treinta minutos. Las muestras son centrifugadas a 6000xg por un minuto. Los 110 µl son colocados en el portaobjeto de vidrio que contiene el microarreglo comercial, el cual es cubierto con otro portaobjeto de vidrio vacío. Esta pareja de portaobjetos es sellada utilizando una cámara de hibridación de microarreglos, para la posterior hibridación a 65 C durante 40 horas en un horno de hibridación con rotación vertical a 20rpm. 48
72 Transcurrido este tiempo las cámaras se desensamblan y los portaobjetos son lavados con la solución amortiguadora 1 (SSPE 0,5X, N-Lauroilsarcosina al 0,005%) por 5 minutos a temperatura ambiente, seguido por un lavado con la solución amortiguadora 2 (SSPE 0,1X, N-Lauroilsarcosina al 0,005%) por 5 minutos a 37 C (solución precalentada a 37 C). Finalmente los portaobjetos son lavados con acetonitrilo durante 10 segundos y luego con una solución estabilizadora (Agilent) por 30 segundos a temperatura ambiente, y retirados lentamente para evitar la formación de burbujas. Luego los portaobjetos se secan completamente, y éstos son escaneados utilizando un Escáner GenePix 4100A y el programa GenePix Pro 5.1, para posteriormente calcular las intensidades de fluorescencia de cada uno de los fluoróforos en cada sonda analizada presente en la plataforma de microarreglos de DNA comercial Análisis e identificación de los cambios en el número de copias en el genoma La imagen de cada uno de los microarreglos obtenida desde el escáner, es analizada con el programa GenePix Pro 5.1, el cual realiza un reconocimiento de cada uno de los pixeles que contiene las sondas, y asigna la localización genómica correspondiente de acuerdo al diseño del microarreglo (diseño ID ). Este programa calcula las medianas de las intensidades de fluorescencias de cada fluoróforo (Cy3-DNA tumor y Cy5-DNA referencia) para cada una de las sondas, sustrayendo un valor de background global. Con estos datos se procede a realizar la normalización de las sondas usando el programa CGHnormaliter en la plataforma R. Éste, corrige los artefactos experimentales mediante un algoritmo iterativo, para finalmente entregar el valor del logaritmo en base 2 de la razón de fluorescencia (tumor/referencia) para cada sonda de cada una de las muestras tumorales. Esta operación permite ajustar todos los valores de las sondas del microarreglo en torno al valor 0 y se obtiene una visualización simétrica de estas razones logarítmicas. De esta manera, se obtuvieron un total de sondas normalizadas del 49
73 microarreglo de DNA. En un contexto ideal, debido a la calidad de las muestras hibridadas, el valor 0 significa que ambas muestras (tumor y referencia) presentan cantidades equivalentes de DNA, denominado estado sin cambio, a diferencia de las muestras que presentan cantidades no equivalentes de DNA, en las cuales se obtienen dos nuevos estados. Uno de éstos es la pérdida o deleción de una copia que es graficada como el Log2 de la razón 1/2 cuyo valor ideal es -1; o bien duplicación o ganancia de una copia que es graficada como el Log2 de la razón 3/2 cuyo valor ideal es +0,58. A continuación, los datos de los logaritmos en base 2 de cada sonda que tiene asignada una localización cromosómica, son ingresados y visualizados en el programa computacional Agilent Genomic Workbench En este programa se realiza la identificación de las regiones genómicas que presentan alteraciones en el número de copias de los cromosomas (CNA). Para esto, el programa contiene algoritmos que permiten identificar las regiones genómicas alteradas en múltiples pares de muestra/referencia. Estos cambios en el número de copias de los cromosomas, ya sea que correspondan a deleciones o ganancias, son identificados con el algoritmo ADM-1 (Aberration Detection Method-1). Este algoritmo detecta intervalos cromosómicos con cambios en el número de copias, con un filtro de la razón logarítmica en base 2 de +0,2 para las ganancias y -0,2 para las deleciones y un intervalo mínimo de 5 sondas con alteración en el número de copias. El algoritmo ADM-1 identifica los intervalos aberrantes o alterados en una muestra dada, basándose en un puntaje estadístico que representa la desviación del promedio de las razones en Log2 normalizadas desde el valor cero esperado. Además, este programa permite graficar a lo largo de cada cromosoma todas las sondas evaluadas en el microarreglo, de manera de detectar y analizar visualmente las regiones genómicas que contienen sondas que presentan variaciones en la hibridación del DNA del tumor con respecto al DNA de referencia., y que refleja alteraciones genómicas en las muestras 50
74 ADM-1 es definido como el promedio de las razones de Log2 normalizadas de todas las sondas en el intervalo genómico multiplicado por la raíz cuadrada del número de sondas que componen el intervalo. Esto representa la desviación del promedio de las razones de Log2 normalizadas desde el valor esperado cero. De esta manera, este algoritmo entrega el puntaje estadístico más alto que encuentra. El gráfico de frecuencia de los cambios en el número de copias del DNA se construye calculando la frecuencia de cada sonda, según la proporción de tumores que portan tal sonda con alteración. El agrupamiento jerárquico no supervisado de los tumores TNBC se realiza con los genes contenidos en las regiones genómicas de pérdida y de ganancia. Pero, de las regiones genómicas de ganancia se seleccionan los genes con el 100% de las sondas dirigidas al gen, con aumento en su número de copias. La medida de distancia usada corresponde a la distancia de Manhattan, con ligamiento completo (complete linkage). Para cada subgrupo de tumores TNBC definidos por la expresión de los marcadores basales y la expresión/localización de BRCA1, se compara el número de regiones de ganancia o deleción, y de los genes contenidos en estas regiones genómicas con alteración contra el resto de las muestras con el fin de identificar las alteraciones en el número de copias específicas de cada subgrupo. Las diferencias entre los subgrupos se evalúan mediante una prueba exacta de Fisher. El valor de p resultante se ajusta con el método de la tasa de falsos descubrimientos (False Discovery Rate; FDR) para corregir la hipótesis múltiple (Benjamini y Hochberg, 1995). Los genes con un valor de p <0.05 y FDR <0.05 se asignan a un subgrupo. Por otra parte, la sobrevida total se calcula desde la fecha del diagnóstico del tumor primario a la fecha de la muerte o del censo de cada paciente. El método de Kaplan-Meier se usa para estimar y construir la curva de sobrevida total. Los análisis estadísticos y la construcción de los gráficos se realizan usando Graph Pad Prism 5.0 y la plataforma de R 51
75 4. RESULTADOS Este estudio se basó en el análisis inmunofenotípico y genómico de 48 tumores de cáncer de mama triple negativo (TNBC) provenientes de 46 pacientes chilenas. Las principales características clínicas e histopatológicas de cada paciente y el tumor respectivo se indican en la tabla 1, en la sección de Métodos. El promedio de la edad de diagnóstico de las pacientes que formaron este grupo fue de 47 años (rango de edad: años). Según, la edad de diagnóstico del cáncer de mama se encontró que un 32,6% (15/46) lo presentó a una edad muy temprana o premenopáusica ( 40 años), un 30,4% (14/46) a una edad temprana o peri menopáusica (entre los años) y un 36,9% (17/46) a una edad tardía o postmenopáusica ( 51 años) (tabla 2). Esto revela que las tres edades de aparición del cáncer de mama se identifican en este grupo de tumores TNBC. Sin embargo, principalmente las pacientes fueron diagnosticadas antes de los 50 años de edad (63% (29/46)). Así, este grupo de cáncer de mama triple negativo está constituido por pacientes con una temprana edad de aparición del cáncer, característico de los tumores TNBC (Tischkowitz y Foulkes., 2006). Por otra parte, se ha descrito que los tumores de mama triple negativo principalmente corresponden a la histología de carcinoma ductal invasor, pobremente diferenciados (grado III). Los antecedentes de nuestros casos muestran que el 90% (43/48) de los tumores fueron carcinoma ductal invasor. De los cuales, un 77% (33/48) corresponden al tipo histológico no especial (nsp) y un 23% (10/48) a carcinoma ductal invasor tipo especial, según las descripciones del anátomopatólogo del hospital respectivo (tabla 2). Se ha descrito, que los tipos histológicos especiales representan menos del 1% de los casos de cáncer de mama en general y que algunos de estos frecuentemente presentan un fenotipo triple negativo. A este respecto, contamos con carcinomas del tipo medular (n = 4) y tipo metaplásico (n = 3), los más recurrentes dentro de los tumores 52
76 TNBC. Además, de tumores con características especiales denominados: tipo papilar (n = 1), tipo mucinoso (n = 1) y tipo tubular (n = 1), de muy baja frecuencia en nuestro grupo de estudio y en la población mundial con cáncer de mama. En relación al estado de diferenciación del tumor, a partir de los informes entregados desde cada unidad de anatomía-patológica del hospital respectivo, se obtuvo información del grado del tumor para 39 carcinomas ductal invasor (tabla 2). De éstos, un 82% (32/39) fueron grado III (pobremente diferenciados), un 15,3% (6/39) grado II (moderadamente diferenciados) y sólo el tumor tipo tubular (T_TN15) fue grado I, (bien diferenciado). Además, se contabilizaron 4 carcinomas ductales in situ y un carcinoma lobular invasor, que representan un 10% de nuestro grupo de estudio (tabla 2). Así, la alta frecuencia de carcinoma ductal invasor, y predominantemente pobre diferenciados (grado III), representan características frecuentes de los tumores de cáncer de mama triple negativo (Reis-Filho y Tutt, 2008; Carey y cols., 2010). 53
77 Tabla 2: Características clínicas e histológicas de las pacientes y sus respectivos tumores de cáncer de mama triple negativo. N total Pacientes 46 Tumores 48 Promedio edad de diagnóstico 47 años Grupos de edades de diagnóstico (n=46) N de Pacientes % de Pacientes Edad muy temprana ( 40 años) 15 32,6 Edad temprana (41-50 años) 14 30,4 Edad tardía ( 51 años) 17 36,9 Histología del tumor (n=48) N de Tumores % detumores Carcinoma Ductal Invasor Carcinoma Ductal In situ 4 8 Carcinoma Lobular Invasivo 1 2 Tipo histológico (n=43) Carcinoma ductal invasor tipo no especial Carcinoma ductal invasor tipo especial Carcinoma ductal invasor tipo especial (n=10) Medular 4 9,3 Metaplásico 3 7 Mucinoso 1 2,3 Papilar 1 2,3 Tubular 1 2,3 Grado del tumor (n=39) I 1 3 II 6 15 III Los símbolos ( ) y ( ), indican menor que y mayor que respectivamente. Los números romanos se refieren al grado de diferenciación de los carcinomas ductal invasor: (I) bien diferenciado, (II) moderadamente diferenciado y (III) pobremente diferenciado. Los porcentajes fueron calculados de acuerdo al n total, indicado entre paréntesis según corresponde. 54
78 4.1 Definición de los tumores TNBC en tipo Basal y tipo no-basal, por inmunohistoquímica. Para definir los tumores de cáncer de mama triple negativo (TNBC) en tipo Basal, se realizó la detección mediante inmunohistoquímica de los marcadores basales: CK5, CK14 y de EGFR. Un tumor TNBC fue definido tipo Basal si expresó la CK5 y/o la CK14 y/o EGFR. En cambio, los tumores sin expresión de estas proteínas (CK5-/CK14-/EGFR-) fueron definidos como tipo no-basal que derivan del panel definido por Nielsen y cols., (2004). El análisis de cada marcador tipo Basal en los 48 tumores TNBC, por inmunohistoquímica, reveló que un 65% (31/48) de los casos expresa la CK5, un 51% (24/47) expresó la CK14, y un 43% (20/46) fueron positivos para EGFR (tabla 3). La diferencia en el total de los tumores analizados para cada marcador fue producto de la ausencia de células tumorales en los casos T_TN06_2 y T_TN28, que resultaron no informativos para el o los marcadores respectivos (tabla 3, figura 5). Así, con este panel de marcadores proteicos, se encontró que un 77% (37/48) de los tumores TNBC se definieron en el subgrupo tipo Basal por expresar uno o varios de los marcadores basales. En cambio, un 23%(11/48) de los casos fueron clasificados como tipo no-basal, ya que resultaron negativos para la expresión de los marcadores CK5-/CK14-/EGFR- (tabla 3, figura 5). 55
79 Tabla 3: Expresión de los marcadores tipo Basal, citoqueratinas 5, 14 y de EGFR, en los tumores de cáncer de mama triple negativo. ID Tumor Histología Tipo Histológico Grado del Edad de CK5 CK14 EGFR Clasificación del del tumor tumor diagnóstico Expresión Puntaje-H Expresión Puntaje-H Expresión Puntaje-H Localización celular Subgrupo T_TN03 Carcinoma ductal invasor no especial II NA tipo no-basal T_TN21_2 Carcinoma ductal invasor Metaplasico III NA tipo no-basal T_TN05 Carcinoma ductal in situ NA NA NA tipo no-basal T_TN06_1 Carcinoma ductal invasor no especial III NA tipo no-basal T_TN09 Carcinoma lobulillar invasor NA NA NA tipo no-basal T_TN19 Carcinoma ductal invasor no especial II NA tipo no-basal T_TN14 Carcinoma ductal invasor no especial III NA tipo no-basal T_TN42 Carcinoma ductal in situ NA NA NA tipo no-basal T_TN46 Carcinoma ductal invasor no especial II NA tipo no-basal T_TN15 Carcinoma ductal invasor Tubular I NA tipo no-basal T5 Carcinoma ductal invasor no especial III NA tipo no-basal T_M156 Carcinoma ductal invasor no especial III NA tipo Basal T42 Carcinoma ductal invasor no especial III NA tipo Basal T_TN48 Carcinoma ductal invasor no especial III NA tipo Basal T_TN49 Carcinoma ductal invasor no especial III NA tipo Basal T_TN04 Carcinoma ductal invasor no especial III NA tipo Basal T11 Carcinoma ductal invasor no especial III NA tipo Basal T_TN50 Carcinoma ductal invasor no especial III NA tipo Basal T_TN38 Carcinoma ductal invasor no especial III membrana tipo Basal T_TN39 Carcinoma ductal invasor no especial III citoplasma/membrana tipo Basal T_TN47 Carcinoma ductal invasor no especial III membrana tipo Basal T_TN43 Carcinoma ductal invasor no especial III citoplasma tipo Basal T_TN08 Carcinoma ductal invasor Papilar NA citoplasma tipo Basal T_TN24 Carcinoma ductal invasor no especial III NA tipo Basal T_TN10 Carcinoma ductal invasor no especial III citoplasma tipo Basal T_TN16 Carcinoma ductal invasor Medular ND citoplasma/membrana tipo Basal T_TN17 Carcinoma ductal invasor Mucinoso III citoplasma/membrana tipo Basal T_TN31 Carcinoma ductal invasor no especial III NA tipo Basal T24 Carcinoma ductal invasor no especial II membrana tipo Basal T_TN11 Carcinoma ductal invasor Medular III membrana tipo Basal T43 Carcinoma ductal invasor no especial III NA tipo Basal T41 Carcinoma ductal invasor no especial III citoplasma/membrana tipo Basal T_TN27 Carcinoma ductal invasor no especial III citoplasma tipo Basal 56
80 Continuación Tipo Histológico del tumor Grado del tumor Edad de diagnóstico CK5 CK14 EGFR Clasificación del ID Tumor Histología Expresión Puntaje-H Expresión Puntaje-H Expresión Puntaje-H Localización celular Subgrupo T_TN28 Carcinoma ductal in situ NA NA NA NA NA tipo Basal T39 Carcinoma ductal invasor no especial III citoplasma tipo Basal T_TN01 Carcinoma ductal invasor Medular ND membrana tipo Basal T_TN06_2 Carcinoma ductal in situ NA NA NA NA NA NA NA tipo Basal T_TN34 Carcinoma ductal invasor no especial III membrana tipo Basal T_TN07 Carcinoma ductal invasor Medular ND citoplasma/membrana tipo Basal T_TN20 Carcinoma ductal invasor no especial II citoplasma/membrana tipo Basal T_TN45 Carcinoma ductal invasor no especial III citoplasma/membrana tipo Basal T_P4 Carcinoma ductal invasor no especial III NA tipo Basal T_TN51 Carcinoma ductal invasor no especial III citoplasma/membrana tipo Basal T_TN44 Carcinoma ductal invasor no especial III citoplasma tipo Basal T_TN02 Carcinoma ductal invasor no especial III membrana tipo Basal T_TN32 Carcinoma ductal invasor no especial III citoplasma/membrana tipo Basal T_TN12 Carcinoma ductal invasor Metaplasico II citoplasma/membrana tipo Basal T_TN21_1 Carcinoma ductal invasor Metaplasico III citoplasma/membrana tipo Basal En la tabla se indica NA: No aplica, ND: No definido. Además, el grado de diferenciación de los carcinomas ductal invasor: (I) bien diferenciado, (II) moderadamente diferenciado y (III) pobremente diferenciado. La intensidad de tinción de cada marcador proteico fue estimada usando los valores: (-): negativo, (1+): débil intensidad, (2+): moderada intensidad, y (3+) fuerte intensidad. 57


81 A). Estructuras de la glándula mamaria normal. B). Tumores TNBC/tipo Basal 58
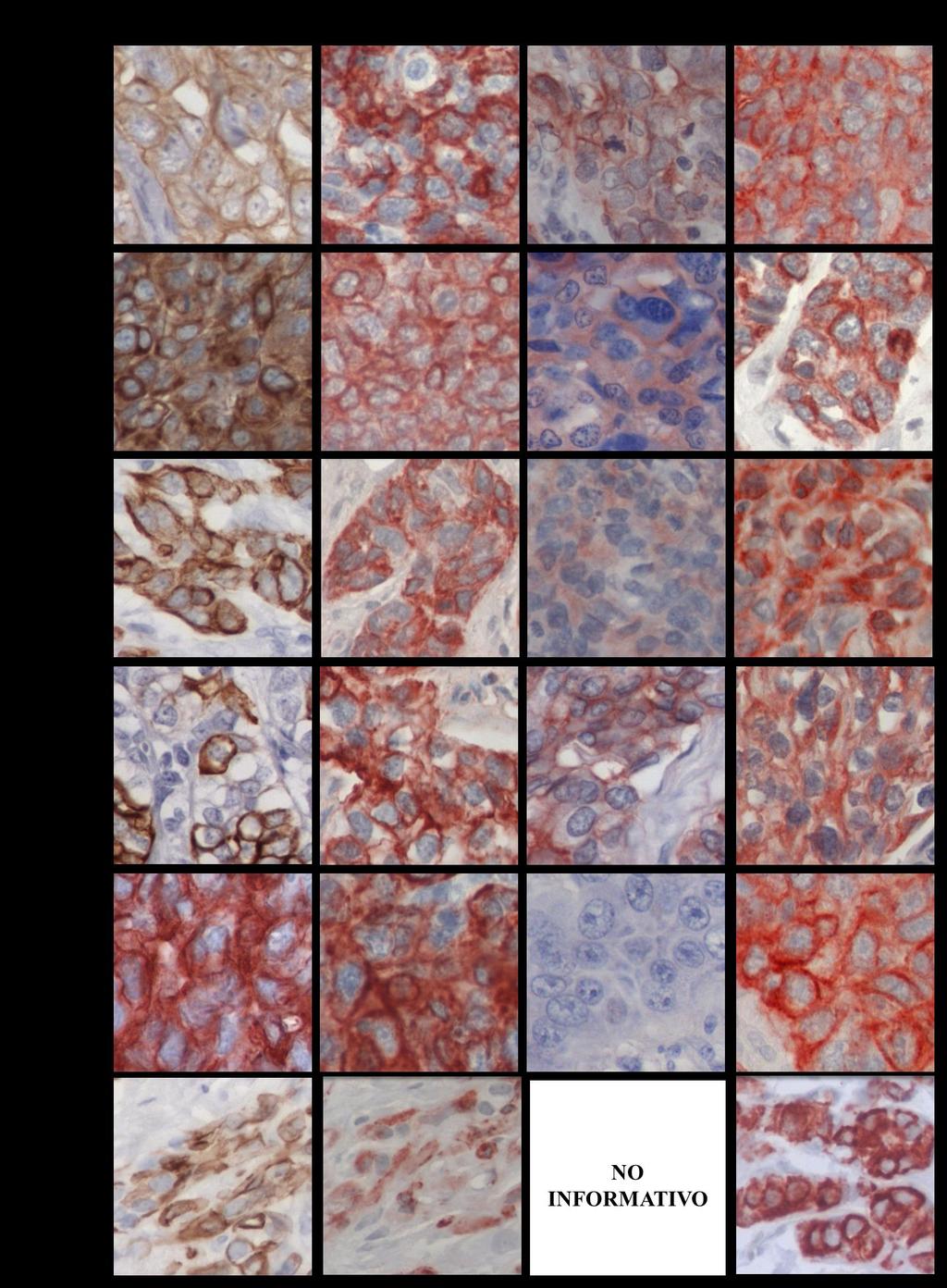
82 59
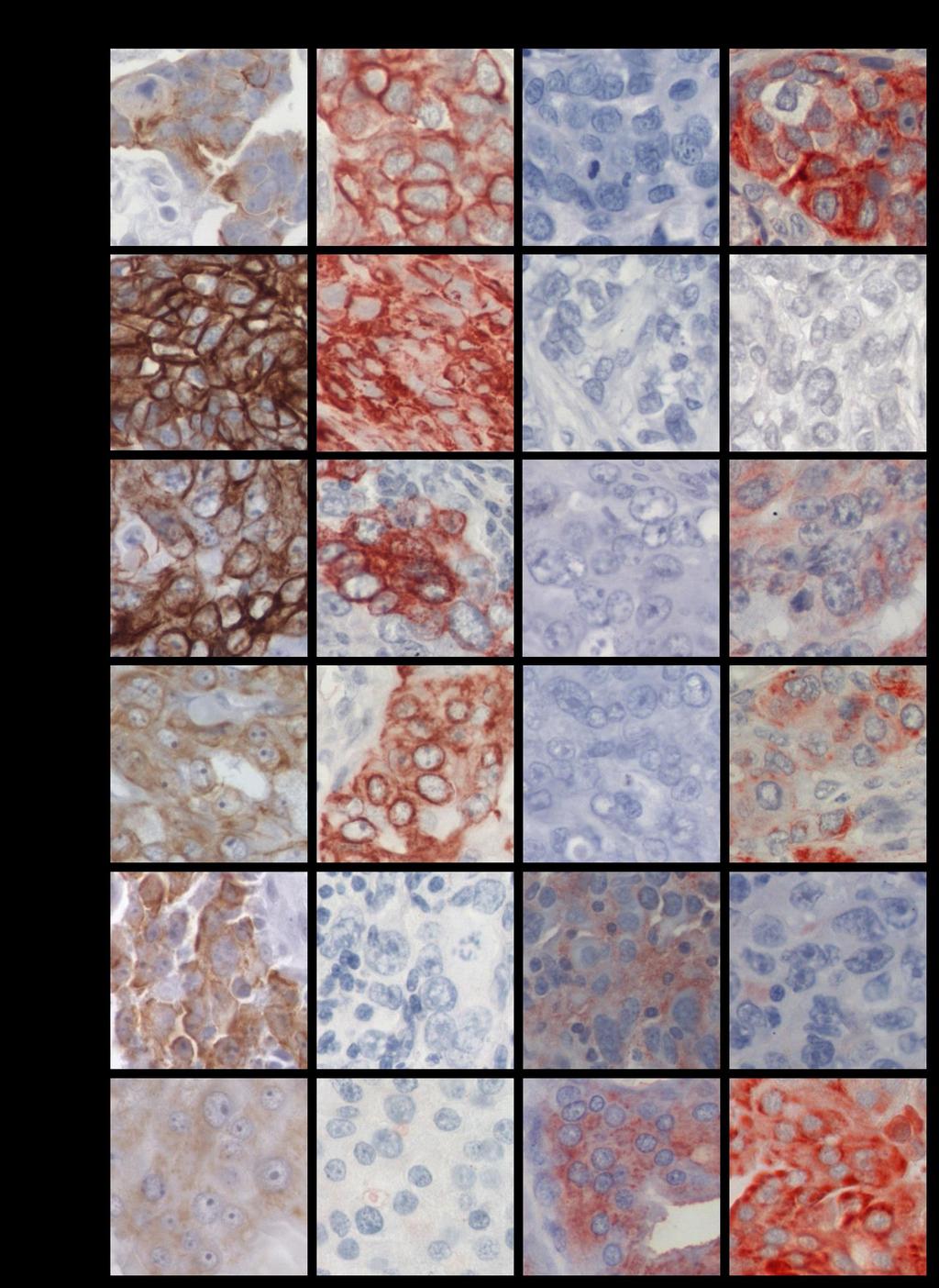
83 60
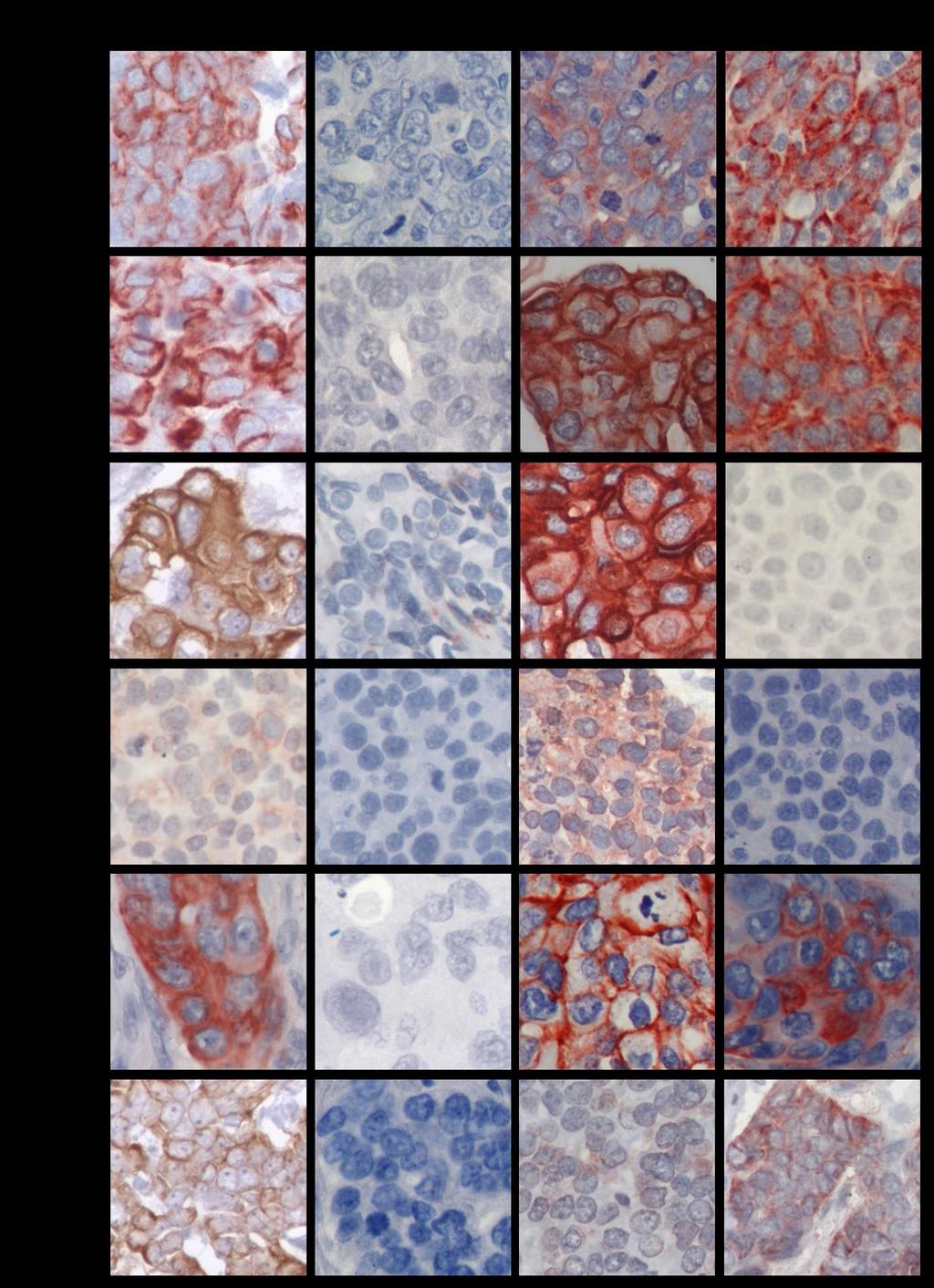
84 61
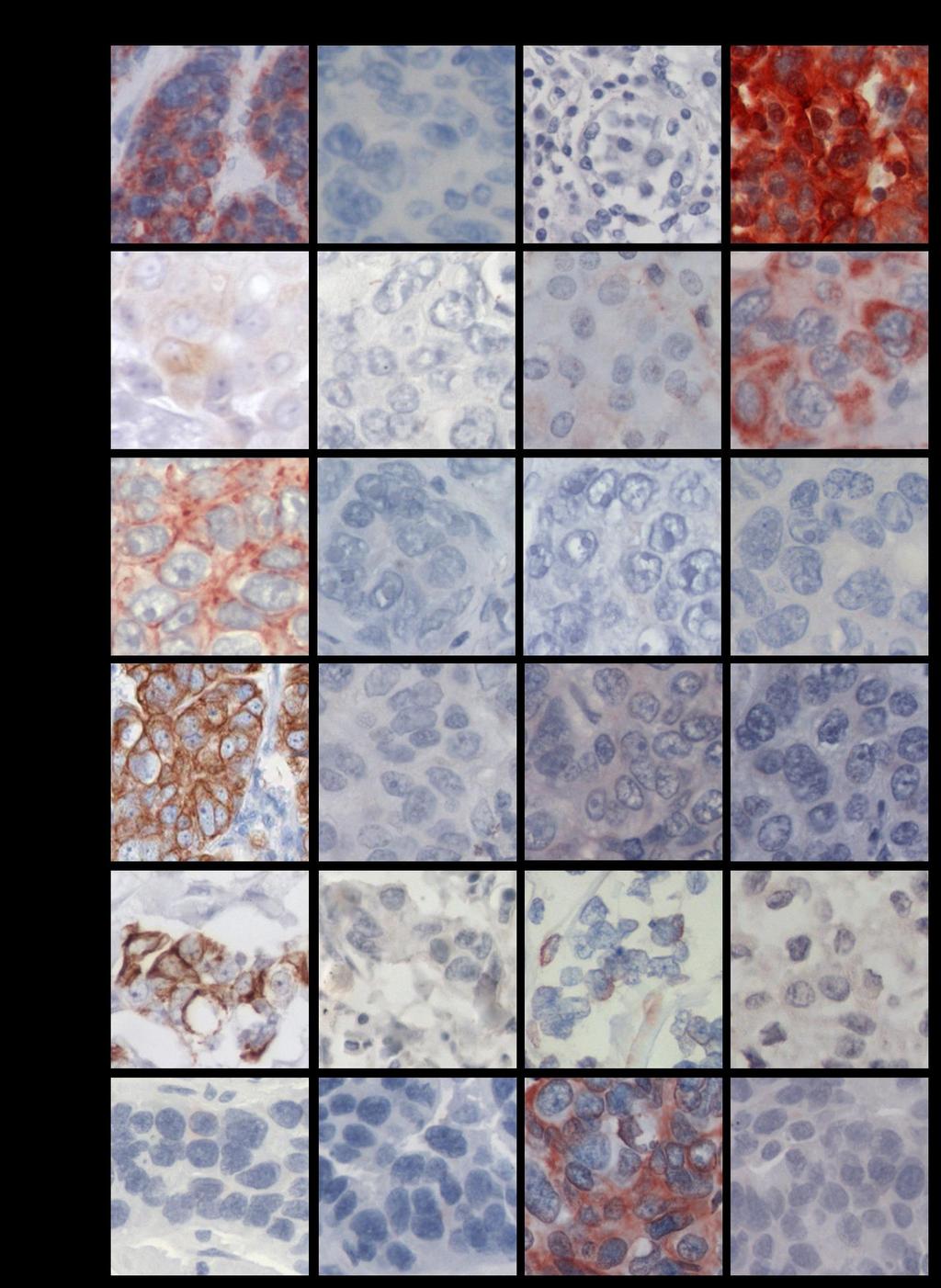
85 62
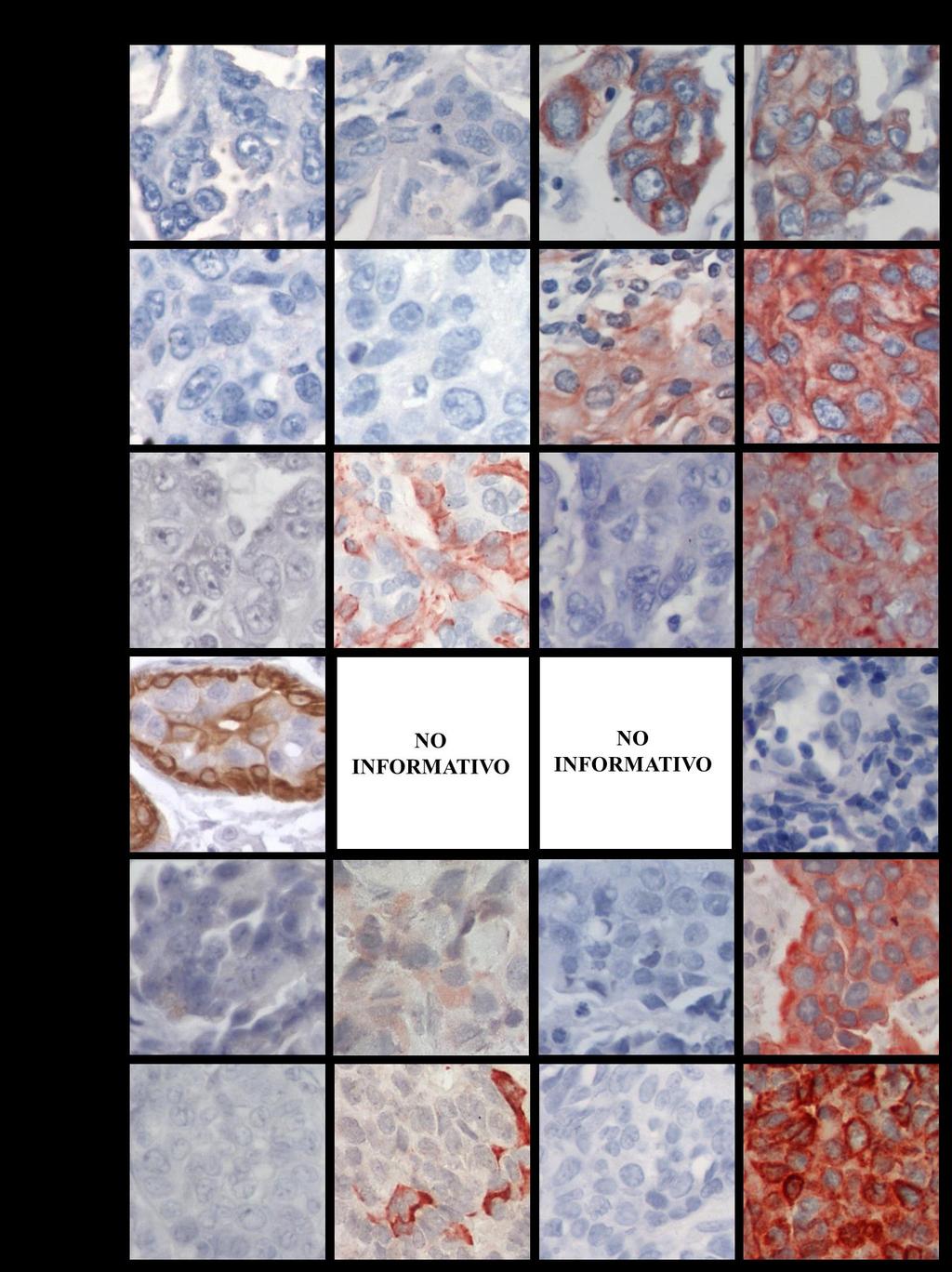
86 63
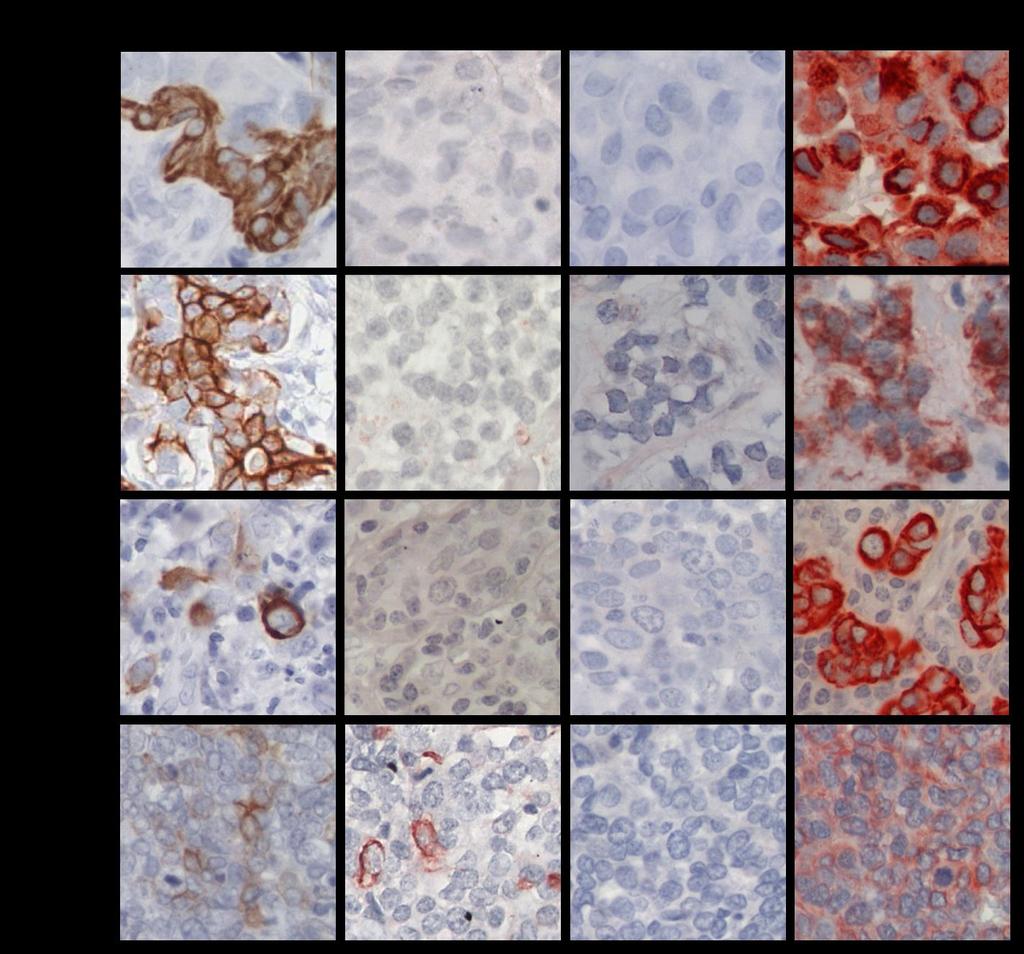
87 64

88 C). Tumores TNBC/tipo no-basal 65

89 Figura 5: Expresión de las citoqueratinas 5, 14, y EGFR en los tumores TNBC. Al costado izquierdo de la figura se indica el tumor TNBC y en la parte superior se indica la proteína correspondiente a la imagen. En la figura B. se indican todos los tumores que fueron definidos como TNBC/tipo Basal. C: Tumores TNBC definidos como tipo no-basal. Los núcleos de las células tumorales se observan de color celeste, el citoplasma está marcado de color rojo. Fotos obtenidas con un objetivo 40X. 66
90 Esto resultados indican que mediante la técnica de inmunohistoquímica, los tumores TNBC frecuentemente presentan un fenotipo tipo Basal, sugiriendo que presentan un perfil de expresión génico similar. Por el contrario, los tumores TNBC/tipo no-basal constituyen un grupo diferente, sugiriendo un perfil de expresión génico distinto a los clasificados en tipo Basal. Este resultado es similar a lo descrito en la literatura, en los cuales se indica que no todos los tumores TNBC corresponden al grupo tipo Basal, revelando la heterogeneidad de los tumores de cáncer de mama triple negativo. Además, demuestra que es insuficiente usar solo la ausencia de expresión del receptor de estrógeno, del receptor de progesterona y de HER2 con el fin de clasificar los tumores tipo Basal como ha sido propuesto en otro trabajo (Cheang y cols., 2008). Principalmente, los tumores TNBC/tipo Basal expresaron conjuntamente más de un marcador proteico en el tejido tumoral. Asi, un 72,9% (27/37) de los casos expresaron tres o dos marcadores basales, sugiriendo que es frecuente la co-expresión de estas proteínas entre los tumores TNBC. Se obtuvieron tres grupos de tumores TNBC/tipo Basal. El 30,5% (11/36) de los casos, expresaron los tres marcadores CK5+/CK14+/EGFR+, un 44,4% (16/36) expresaron solo dos marcadores tipo Basal (CK5+/CK14+= 62,5% (10/16); CK5+/EGFR+ = 37,5% (6/16)) y finalmente, un 27% (10/37) expresó solo un marcador tipo Basal. En este último grupo, la expresión de la CK5+ o CK14+, se observó en aproximadamente en un 5% de las células tumorales del área del tumor. Esto sugiere, que una población focal de células tumorales mantiene la expresión de la citoqueratina correspondiente, mientras que las células restantes pierden la expresión de estas citoqueratina a medida que el tumor se va desdiferenciando. En cambio, los TNBC/tipo Basal con solo expresión de EGFR mostraron expresión principalmente en la membrana entre un 10-70% de las células tumorales, con fuerte intensidad en los tumores respectivo (tabla 3). Esto sugiere que los tumores positivos para EGFR se encuentran en un estado 67
91 con aumento de la proliferación celular. Por su parte, aquellos tumores con expresión conjunta de los tres o dos marcadores basales mostraron un mayor número de células positivas con tinción y una distribución heterogénea de cada marcador. Al analizar la expresión de cada marcador basal en el grupo definido TNBC/tipo Basal, el que mayormente los definió fue la expresión de la CK5 en un 83,7% (31/37) de los casos. A diferencia de la expresión de la CK14 que fue de un 65% (24/37) y de EGFR en un 54% (20/37) de los tumores. Esto indica que la expresión de la CK5 es muy importante para definir los casos tipo Basal en el conjunto de tumores TNBC. Respecto a la expresión de las citoqueratinas basales, se ha descrito que la CK5 y la CK14 se aparean para formar los filamentos intermedios de las células basal/mioepitelial de la mama normal, y su patrón de expresión se mantiene durante el proceso de transformación, observándose en las células del tumor (Moll y cols., 1982). De acuerdo a estas evidencias, se esperaba obtener similar proporción de tumores y de sus células tumorales con expresión de ambas citoqueratinas. Sin embargo, los resultados mostraron que un 57% (21/37) de los TNBC/tipo Basal fueron positivos para ambas citoqueratinas (CK5+/CK14+) conjuntamente, con una fuerte intensidad de tinción (3+), y clara localización citoplasmática. Incluso, existe una significativa correlación (valor-p de 0,0008) respecto a la expresión de ambas citoqueratinas en los tumores TNBC/tipo Basal, sugiriendo co-expresión de CK5 y CK14 en estos tumores. Pero, un 24% (9/37) de los casos CK5-positivos no expresa la CK14, la cual incluso se expresa en un menor porcentaje de células tumorales, sugiriendo que hay una pérdida de expresión de la CK14 en estos tumores. Por lo tanto, muchos de los tumores TNBC/tipo Basal presentan formación de los filamentos intermedios utilizando el par de CK5/CK14, las cuales se co-expresan en la misma célula. Sin embargo, en las células malignas principalmente se mantiene la expresión de la CK5 y la CK14 va 68
92 perdiendo su expresión. Estos resultados sugieren que existe una pérdida de expresión de la CK14 o bien una desregulación de la expresión de esta citoqueratina en las células tumorales de los tumores TNBC/tipo Basal. Por otra parte, el análisis de los tumores positivos para EGFR mostró diferentes localizaciones subcelulares de este receptor. Un 30% (6/20) de los casos lo expresó en la membrana de las células tumorales, un 45% (9/20) lo expresó tanto en el citoplasma y membrana, en cambio en un 25% (5/20) se localizó solo en el citoplasma de las células tumorales. La localización de EGFR en la membrana y el citoplasma/membrana, sugiere que en estos casos EGFR se encuentra funcional, produciendo una señalización de proliferación. La sobre-expresión del receptor de EGF, no es específico de las células basal/mioepitelial, pero en este subtipo molecular se ha encontrado expresado en un 60% de los tumores descritos por Perou y cols., (2000) y fue incluido como un marcador para definir tumores tipo Basal por Nielsen y cols., (2004). A este respecto, identificamos tumores TNBC/tipo Basal con aumento de expresión de EGFR en conjunto con la CK5. Independiente de la localización citoplasmática o de membrana se encontró una asociación significativa de estas dos proteínas en estos tumores (valor-p fue de 0,0127). Esta asociación, sugiere que la CK5 y EGFR fueron co-expresados en las células tumorales de los tumores TNBC/tipo Basal. Aún más, no se identificaron tumores con la expresión conjunta de CK14 y de EGFR, solo esto se observó en los tumores con expresión de los tres marcadores (CK5+/CK14+/EGFR+). Sin embargo, en estos casos no podemos demostrar que se encuentren colocalizando en la misma célula tumoral. Con este panel de tres marcadores tipo Basal podemos demostrar que los tumores TNBC/tipo Basal presentan patrones de expresión heterogéneos tanto inter-tumor como intratumor, siendo predominante la expresión de la CK5. Además, sugiere dos posibilidades de acuerdo 69
93 a la literatura: 1) que existen diferentes poblaciones de células tumorales, indicando que las células iniciales corresponden a CK5+/CK14+, pero estas con el tiempo van perdiendo su expresión, siendo más susceptible la pérdida de expresión de la CK14+. 2) que las células tumorales van desde un estado diferenciado a desdiferenciado. Así, a medida que los tumores se van desdiferenciando van perdiendo sus características iniciales que se ve reflejado por una pérdida del patrón de expresión de las citoqueratinas iniciales. 4.2 Análisis de la expresión y localización de la proteína BRCA1 en la glándula mamaria normal y los tumores TNBC. La expresión de la proteína BRCA1 fue evaluada en tres tejidos provenientes de mama normal y en los 48 tumores TNBC, utilizando un anticuerpo monoclonal dirigido al N-terminal de esta proteína. En los tejidos de mama normal, se observó expresión de la proteína BRCA1 en el núcleo de las células epiteliales que forman los ductos y lóbulos (figura 6). Principalmente, con moderada y fuerte intensidad de tinción. Sin embargo, en uno de estos tejidos se identificaron algunas estructuras ductales cuyos núcleos de las células luminales, mostraron una intensidad moderada (2+) a diferencia de las células basal/mioepitelial con débil intensidad (1+). Estas diferencias no fueron observadas en las estructuras lobulares. En los tres tejidos, no se obtuvo expresión de BRCA1 en el citoplasma o membrana. Los resultados revelan que BRCA1 se expresa en el núcleo de las células luminal y basal/mioepitelial de la glándula mamaria normal, aunque en estas últimas células se puede observar una menor intensidad de tinción de BRCA1 en algunos ductos de la mama. En la figura 6 se muestran imágenes de la inmunotinción de BRCA1 en estructuras de la glándula mamaria normal. En contraste, a los tejidos de mama normal, los tumores TNBC malignos, mostraron intensidades de tinción variables, alterada localización subcelular y una heterogénea tinción 70
94 nuclear tanto inter-tumor como intra-tumor. Inmunoreactividad al anticuerpo anti-brca1 se detectó en un 91,7% (44/48) de los TNBC. Por el contrario, en un 8,3% (4/48) de los casos se obtuvo una completa ausencia de expresión de BRCA1, en las células tumorales. Esto indica que los tumores TNBC frecuentemente expresan la proteína BRCA1, aunque se identifican tumores con ausente expresión de esta proteína (Figura 7). Sin embargo, como se explicó en la introducción, BRCA1, es una proteína que predominantemente se localiza en el núcleo, ya que participa en la reparación del DNA por quiebre en la doble hebra, por lo tanto, su expresión en esta ubicación celular es requerida para su normal funcionamiento. A este respecto, los niveles relativos de expresión o la intensidad de la tinción, el porcentaje de células tumorales positivas, y una incorrecta localización en el citoplasma, dan cuenta de la ausente o reducida expresión de esta proteína que se traduce en una pérdida de función de BRCA1 en las células tumorales. En este contexto, evaluamos la expresión de BRCA1, según la localización subcelular, la intensidad de tinción y el porcentaje de células positivas en nuestro grupo de tumores. De los tumores TNBC positivos para BRCA1, un 47,7% (21/44) expresó esta proteína en el núcleo de las células tumorales. De estos, 10 tumores expresaron BRCA1 con intensidades moderadas y fuertes, en cambio en 11 tumores la expresión fue débil en el núcleo. Por otra parte, un 4,5% (2/44) expresó BRCA1 solo en el citoplasma y un 47,7% (21/44) la expresó tanto en el núcleo como en el citoplasma de las células tumorales. Esto indica, que si bien en los tumores TNBC se expresa BRCA1, se pueden observar diferentes patrones de expresión de esta proteína, siendo predominante identificar tumores TNBC con una alterada localización subcelular. Esto nos revela que la expresión de BRCA1 en los tumores TNBC es heterogénea, en relación a la intensidad de tinción y la localización subcelular. 71
.")

95 Figura 6: Imágenes de la inmunohistoquímica de BRCA1 en tejidos de mama normal, provenientes de reducciones mamarias. En la figura A: se muestra una estructura lobular, sin exposición al anticuerpo BRCA1 (control negativo). B: Estructura lobular, donde se observan acinos cuyas células epiteliales presentaron inmunotinción de BRCA1 en el núcleo. C: Se muestra una estructura ductal, cuyas células luminales y basales/mioepteliales expresaron BRCA1 en el núcleo, con fuerte intensidad de tinción. D: Imagen de una estructura ductal, con mayor intensidad de tinción en las células luminales que las células basales/mioepiteliales. Fotos obtenidas con un objetivo 40X.. 72

96 A). Tumores TNBC con tinción nuclear de BRCA1. 73
.")
97 B). Tumores TNBC con ausente o débil tinción de BRCA1 en el núcleo. B.1). Ausente tinción 74

98 B.2). Débil tinción 75
.")
99 C). Tumores TNBC con tinción de BRCA1 nuclear y citoplasmática. C.1). Débil tinción 76
 Expresión en el citoplasma con moderada/fuerte intensidad de tinción. B) Ausente o débil expresión nuclear de BRCA1.")
100 C.2). Moderada/fuerte tinción Figura 7: Imágenes de la expresión y localización de BRCA1 en los tumores TNBC. A) Expresión en el citoplasma con moderada/fuerte intensidad de tinción. B) Ausente o débil expresión nuclear de BRCA1. C) tumores TNBC con expresión de BRCA1 en núcleo y citoplasma. Fotos obtenidas con un objetivo 40X. 77
101 Debido a los diferentes patrones de expresión de la expresión de BRCA1, y la baja proporción de tumores TNBC con ausente expresión de BRCA1 se agruparon en tres grupos (Figura 8). Un grupo fue denominado con expresión nuclear de BRCA1 que corresponde a un 20,8% (10/48) de los tumores TNBC. Estos casos mostraron intensidades moderadas (2+) y fuertes (3+) en el núcleo, similar a los tejidos de mama normal, y en un porcentaje mayor al 60% de las células tumorales. El segundo grupo fue llamado con pérdida de expresión nuclear de BRCA1. Este representó un 31,3% (15/48) de los casos e incluye, los 11 tumores con débil (1+) intensidad de tinción nuclear y los 4 tumores con ausencia completa de la expresión de BRCA1. Finalmente, el último grupo fue nombrado con expresión de BRCA1 citoplasmática con/sin expresión nuclear conformado por un 47,9 (23/48) de los casos TNBC. Dentro de este grupo se encuentran los 2 tumores con solo expresión en el citoplasma (T_TN08 y T_TN38), y los 21 tumores con expresión en el núcleo y citoplasma, con variables intensidades de tinción. En este último grupo, se observó que el promedio de células tumorales positivas para BRCA1 en el núcleo fue de un 67% y en el citoplasma fue de un 71%, sugiriendo que una misma célula tumoral expresa BRCA1 en ambos compartimentos subcelulares. Estos resultados revelaron que el 79% de los tumores TNBC presentan una reducida expresión nuclear o una alterada localización de la proteína BRCA1, sugiriendo que la pérdida de expresión de BRCA1 y su alterada localización pueden ser relevantes para la progresión del tumor. 78

102 Figura 8: Gráfico de los tres grupos principales de expresión y localización de la proteína BRCA1. 79
103 4.2.1 Análisis de la expresión de la proteína BRCA1 en los tumores con mutación en BRCA1 o BRCA2 en la línea germinal. Entre los tumores TNBC, se encontraron 7 casos de pacientes portadoras de una mutación en el gen BRCA1 (n = 3) o BRCA2 (n = 4) en la línea germinal. De los tumores con mutación en el gen BRCA1, el caso T_P4, expresa BRCA1 en el 10% de las células tumorales y el tumor T_TN17, expresa débilmente BRCA1 en el 40% de las células tumorales. En cambio, el tumor T24 presentó expresión de BRCA1 en el 40% de las células tumorales en el núcleo, y en el 80% de las células en el citoplasma. Para este último tumor, la intensidad de tinción fue débil (1+) en ambos compartimentos subcelulares. Por lo tanto, existe la posibilidad que el alelo normal de BRCA1 este expresando una isoforma que carece del exón 11 de BRCA1, donde se localiza la señal de importe nuclear, de esta forma podríamos estar observando BRCA1 en el citoplasma celular. En estos tres tumores de pacientes portadoras de una mutación en el gen BRCA1 en la línea germinal, se observó un bajo porcentaje de células tumorales con débil expresión de la proteína BRCA1. Por lo tanto, existe una menor fracción de células tumorales con expresión de BRCA1, sugiriendo que somáticamente existen células que no han sufrido una completa pérdida de heterozigocidad (LOH). Este fenómeno, sugiere que un gen supresor de tumor, como BRCA1, debe perder ambos alelos con el fin de obtener una completa pérdida de función. Por otra parte, la expresión de BRCA1 en el citoplasma de las células tumorales sugiere que se expresa una isoforma, que carece del exón 11 de BRCA1, donde se localiza la señal de importe nuclear, de esta forma podríamos estar observando BRCA1 en el citoplasma celular. En relación a los tumores de pacientes portadoras de mutación en el gen BRCA2, los casos T_M156 y T_TN10 expresaron BRCA1 solo en el núcleo, con una fuerte intensidad. En cambio, 80
104 los casos T_TN39 y T5 la expresaron en ambos compartimentos celulares. Así, se observa una variabilidad en la expresión de la proteína BRCA1 en estos tumores, lo cual sugiere que aunque BRCA2 este mutado, hay casos en los cuales BRCA1 se ve afectado, por su alterada localización celular Expresión y localización de BRCA1, según la clasificación tipo Basal y tipo no- Basal de los tumores TNBC. Existe relación entre la pérdida de expresión de BRCA1 o su alterada localización citoplasmática con la clasificación tipo Basal o tipo no-basal de los tumores TNBC? Se ha sugerido que los tumores de mama definidos como tipo Basal presentan una pérdida de función de BRCA1, ya que aproximadamente un 80% de los tumores provenientes de portadoras de mutación en el gen BRCA1 en la línea germinal, se sobreponen con el fenotipo tipo Basal (Sorlie y cols., 2001). Sin embargo, esta correlación en relación a la expresión de la proteína BRCA1 o su localización subcelular no ha sido ampliamente evaluada. En este estudio, se encontró que un 77% (37/48) de los tumores corresponden al subgrupo TNBC/tipo Basal y un 23% (11/48) al subgrupo TNBC/tipo no-basal. Por otra parte, determinamos los niveles relativos de la expresión de la proteína BRCA1 en el núcleo y citoplasma, de manera independiente (Figura 9). Esto fue representado con el puntaje-h que corresponde al producto de la intensidad de tinción por el porcentaje de células positivas de BRCA1. En relación a la expresión de BRCA1 en el núcleo se usó la mediana del puntaje-h =100, como el valor umbral para definir los tumores con ausente o reducida expresión de BRCA1 (pérdida de expresión de BRCA1 nuclear), clasificados por un puntaje-h 100 y los tumores con expresión de BRCA1 en el núcleo, definidos por un puntaje-h >100. Así, un 58,3% (28/48) de los tumores presentaron un puntaje-h 100, representando el grupo con reducida o ausente expresión 81
105 de BRCA1 nuclear. En este grupo de tumores, la mediana del porcentaje de células positivas para BRCA1 fue de un 50%, indicando que existe pérdida de expresión de BRCA1 nuclear en el 50% restante de las células tumorales. En contraste, un 41,7% (20/48) de los casos obtuvo un puntaje- H >100, los cuales fueron determinados con expresión de BRCA1 en el núcleo. Principalmente, este grupo quedo conformado por tumores con una moderada o fuerte intensidad de tinción nuclear de BRCA1. La mediana del porcentaje de células positivas para BRCA1 fue de 80%, indicando que en estos tumores BRCA1 se expresa en la mayoría de las células tumorales. Del grupo de tumores con una ausente o reducida expresión de BRCA1 nuclear ( puntaje- H 100), el 89% (25/28) corresponden a TNBC/tipo Basal y solo 3 tumores (11%) corresponden a tumores TNBC/tipo no-basal. Este resultado sugiere que existe una tendencia de que los tumores tipo Basal, presenten una pérdida de expresión de BRCA1 como ha sido postulado para los tumores con mutación en el gen BRCA1 en la línea germinal. Aún más, los 3 tumores con mutación en la línea germinal del gen BRCA1 presentan un fenotipo tipo Basal. En cambio, del grupo de tumores con expresión de BRCA1 nuclear (puntaje-h > 100) el 60% (12/20) fue clasificado TNBC/tipo Basal y un 40% (8/20) corresponden al fenotipo TNBC/tipo no-basal. Esto indica que un 72% (8/11) de los tumores TNBC definidos en tipo no-basal, principalmente expresan la proteína BRCA1 en el núcleo, sugiriendo que en estos tumores BRCA1 se encuentra funcional. Con estos resultados se encontró que existe una asociación significativa entre la ausencia o reducida expresión de BRCA1 y el fenotipo TNBC/tipo Basal, con un valor de p = (Figura 9). Esto demuestra que existe una correlación entre la pérdida de expresión nuclear de BRCA1 y el subtipo TNBC/tipo Basal, sugiriendo que en estos tumores existe pérdida de función de BRCA1. 82
106 Por otra parte, en relación a la expresión citoplasmática de BRCA1 se encontró que un 65% (15/23) corresponden a tumores TNBC/tipo Basal, y un 35% (8/23) fueron clasificados TNBC/tipo no-basal. Así, considerando los tumores con expresión de BRCA1 en el citoplasma, se determinó que la mediana del puntaje-h que representa la expresión de BRCA1 en el citoplasma fue de 80. Utilizando este valor como umbral de expresión de BRCA1 se encontró que un 57% (13/23) de los tumores presentan una disminuida expresión de BRCA1 en el citoplasma (puntaje- H 80). A diferencia del 43% (10/23) restante que muestra expresión de BRCA1 en el citoplasma con un puntaje-h >80. De esta manera, se encontró que no hay correlación entre la expresión citoplasmática de BRCA1 y el fenotipo tipo Basal, cuyo valor de p fue de 0,6850. Esto demuestra que la pérdida de expresión nuclear de BRCA1 y no la expresión en el citoplasma se asocia con el fenotipo tipo Basal. Esto resultados sugieren que los tumores con pérdida de expresión de BRCA1 nuclear pueden conservar un perfil de expresión génica similar, a diferencia de los tumores que presentan BRCA1 en el citoplasma. 83
107 Puntaje-H Expresión de BRCA1 T_TN04 T_TN47 T_TN16 T_TN45 T_TN08 T_TN38 T_P4 T_TN49 T_TN17 T41 T_TN27 T_TN06_2 T24 T_TN06_1 T_TN11 T_TN07 T_TN46 T_TN01 T_TN34 T_TN05 T42 T_TN39 T_TN21_1 T_TN24 T_TN32 T_TN43 T_TN20 T_TN12 T_TN10 T_TN14 T_TN50 T39 T_TN15 T_TN48 T_TN31 T_TN02 T_TN21_2 T_TN19 T43 T_TN03 T_TN44 T_M156 T_TN51 T_TN42 T5 T_TN09 T_TN28 T11 Núcleo Citoplasma tipo Basal tipo no-basal Figura 9: Gráfico de la expresión de BRCA1 en los tumores de cáncer de mama triple negativo. Las barras azules indican la expresión en el núcleo y las barras rojas indican expresión de BRCA1 en el citoplasma. En la parte inferior del gráfico de barras, se muestran en rectángulos naranjos que corresponden a los tumores TNBC/tipo Basal y en azul los casos TNBC/tipo no-basal. La línea verde, corresponde a los tumores con pérdida de expresión de BRCA1 en el núcleo (puntaje-h 100). En cambio, la línea amarilla se refiere a los casos con expresión nuclear de BRCA1 (puntaje-h > 100). 84
108 4.3. Hibridación genómica comparativa en microarreglos de DNA de los tumores de cáncer de mama triple negativo. Una vez caracterizados los tumores TNBC, con respecto al subtipo tipo Basal y tipo no- Basal, y analizado la expresión y localización de la proteína BRCA1, por inmunohistoquímica, se analizó el genoma de los tumores TNBC. Se sabe que BRCA1 participa en la reparación del DNA por recombinación homóloga (Lord and Ashworth, 2012). Esta función celular es muy importante para mantener la estabilidad genómica en las células normales y células tumorales (Lord and Ashworth, 2012). Por lo tanto, se caracterizó el genoma de los 48 tumores TNBC, mediante la técnica de hibridación genómica comparativa en microarreglos de oligonucleótidos de DNA (array-cgh), con el fin de identificar cambios en el número de copias del DNA específicos, en relación a la expresión y localización de BRCA1. Los cambios en el número de copias del DNA o rearreglos genómicos, fueron detectados en los 22 cromosomas autosómicos y el cromosoma X sexual. Este último, sirvió como control interno de amplificación, como se detalla más adelante. Los rearreglos genómicos fueron definidos como: ganancia (aumento en el número de copias de los cromosomas), deleción (pérdida en el número de copias de regiones cromosomales) o neutras, (regiones genómicas inalteradas en su número de copias). Estas regiones genómicas con ganancia o deleción pueden contener potenciales oncogenes o genes supresores de tumor respectivamente, relevantes para la progresión del cáncer de mama triple negativo. 85
109 4.3.1 Porcentaje de sondas con alteración de cada cromosoma, detectado en el microarreglo de oligonucleótido de DNA, entre los tumores TNBC. Los cambios en el número de copias del DNA fueron detectados como intervalos genómicos (rearreglo genómico), compuestos por un número de sondas, en cada muestra TNBC. Los rearreglos genómicos, comprendieron en total sondas con alteración, a través de todos los cromosomas, correspondiendo a un 47,6% (20121/42278) de las sondas normalizadas del microarreglos de DNA. Esto revela que una gran fracción del genoma de los tumores TNBC presenta cambios en el número de copias del DNA somáticos, con ganancia y deleción, sugiriendo inestabilidad genómica de los tumores TNBC. Para cada cromosoma, se determinó el porcentaje de sondas con cambios en el número de copias del DNA (figura 10A). Con mayor porcentaje de sondas con alteración fueron los cromosomas: 17 (99%), 19 (96%), 22 (100%) y el cromosoma X (100%). En cambio, los cromosomas: 4 (16%), 13 (18%) y 18 (19%) resultaron con el menor porcentajes de sondas con alteración, indicando que conservan cantidades equivalentes de DNA genómico entre el tumor y el DNA de referencia. Esto indica que los cromosomas 17, 19 y 22 concentran la mayor cantidad de rearreglos genómicos en los tumores TNBC, posiblemente durante la progresión del tumor. A este respecto, se ha descrito, que los cromosomas 17, 19 y 22, tienen la más alta densidad de genes en el genoma humano, y los cromosomas 4, 13 y 18 presentan la menor densidad de genes, como se muestra en la figura 10B (Mouse Genome Sequencing Consortium, 2002). Además, los cromosomas 17 y 19 contienen varios genes involucrados en el inicio y progresión de diferentes tipos de canceres, como el cáncer de mama (Pan y Epstein, 2010). Entonces, esto sugiere que cromosomas 17 y 19 posiblemente contengan varios sitios de quiebre en su secuencia de DNA, susceptibles a mecanismos que alteran su número de copias del DNA (Przybytkowski y cols., 2014). 86
110 Densidad de CpG (Islotes Mb -1 ) Número de sondas por cromosoma A) Cromosomas % de Sondas con alteración X N Sondas por Cromosoma (Total) N Sondas por cromosoma (alteradas) B) Cromosomas Figura 10: Porcentaje de sondas con cambio en el número de copias en cada cromosoma y la densidad de genes descrita para cada uno de ellos. A) Gráfico de las sondas con alteración en cada cromosoma, entre los tumores TNBC. En el eje Y se muestra el número de sondas normalizadas por cromosoma y en el eje X se indican los cromosomas y el porcentaje de sondas alteradas. Las barras azules corresponden al número de sondas, en cada cromosoma, que contiene el microarreglos de DNA 4X44 previo a la detección de las regiones genómicas con alteración mediante el algoritmo ADM-1. Las barras rojas indica el número de sondas con cambios en su número de copias del DNA, en cada cromosoma. ADM-1: algoritmo que define los intervalos con cambio en su número de copias en cada muestra TNBC. B) Densidad de los sitios CpG en los 24 cromosomas del genoma humano y del genoma de ratón. Los diamantes rojos corresponden al genoma humano y los diamantes azules al genoma de ratón. La alta densidad de los sitios CpG se correlaciona con la alta densidad de genes codificantes en cada uno de los cromosomas. Figura extraída del artículo de Mouse Genome Sequencing Consortium. Nature. 2002; 420(6915):
111 4.3.2 Control interno de ganancia, cromosoma X, en los tumores TNBC. La metodología empleada incluyó un DNA de referencia que corresponde a un pool de DNA genómico proveniente de linfocitos de 10 hombres, el que fue co-hibridado con el DNA genómico tumoral proveniente de mujeres. Esta estrategia, permitió usar el cromosoma X como control interno de ganancia o amplificación ya que las mujeres portan dos cromosomas X y el hombre solo una copia. Así, para el cromosoma X, se obtuvo 2 copias por 1 copia proveniente del pool de DNA genómico de hombres o DNA de referencia. La razón de log2 (DNAtumor/DNAreferencia) fue de 1, lo cual se visualizó en la plataforma del software desviado hacia el +1 (figura 11). Un total de 42 tumores presentaron ganancia o duplicación del cromosoma X (desviado hacia +1), que correspondió a lo esperado. Sin embargo, se identificaron los casos T_TN27, T_TN51, T_TN09, T11, T42 que presentaron este cromosoma alineado en 0, indicando pérdida de 1 copia del cromosoma X. A su vez, el tumor T_TN08 presentó 2 copias del cromosoma X (desviado hacia +2), como se observa en la figura 11, indicando una amplificación de 2 copias de este cromosoma. Por lo tanto, se obtuvo que un 90% (43/48) de los tumores presentaron amplificación de una copia del cromosoma X, con aproximadamente el 99% de las sondas localizadas en este cromosoma sexual. En cambio, el 10% (5/48) de los tumores restantes presentaron pérdida de una copia del cromosoma X. A este respecto, se ha descrito que el cromosoma X sufre de isodisomía o alteración en su número de copias en los tumores de mama, fenómeno particularmente observado en los tumores tipo Basal (Richardson y cols., 2006). Así, 4 de los 5 tumores con deleción de una copia del cromosoma X corresponden a TNBC/tipo Basal, indicando que un 11% (4/37) de los casos TNBC/tipo Basal tienen pérdida de una copia del cromosoma X. Estos resultados indican que la calidad de la muestra fue adecuada y la estrategia experimental fue correctamente aplicada. 88
112 T_TN27 T_TN21_ T_TN08 Centrado en 0 Desviado al +1 Desviado al +2 Figura 11: Visualización del cromosoma X en los tumores TNBC. En la figura se muestra el cromosoma X de 3 tumores representativos. El caso T_TN27 se encuentra centrado en 0, indicando pérdida de una copia del cromosoma X. El caso T_TN21_1, muestra ganancia de una copia y el T_TN08 presenta ganancia de 2 copias del cromosoma X. Figuras obtenidas del programa Agilent Genomic Workbench
113 4.3.3 Análisis de las sondas que reconocen el gen BRCA1, en el microarreglo de oligonucleótido de DNA. La plataforma del microarreglo de oligonucleótido de DNA, que se usó en este estudio, contiene más de una sonda para reconocer un gen relacionado al desarrollo o progresión del cáncer. A este respecto, BRCA1, un gen supresor de tumor, de alto riesgo a desarrollar cáncer de mama, es reconocido por 4 sondas espaciadas dentro de la plataforma. Se ha descrito que los tumores de mama provenientes de portadoras de mutación en el gen BRCA1 en la línea germinal, frecuentemente pierden el segundo alelo por deleción de una región cromosómica o microdeleciones que involucra el gen BRCA1, fenómeno denominado como pérdida de heterozigocidad (LOH). Por lo tanto, con el fin de determinar si los tumores TNBC presentan deleción de BRCA1 se evaluó la región cromosómica 17q21.31, donde se localiza este gen. Mediante el algoritmo ADM-1 y un filtro de 5 sondas consecutivas con alteración, no se detectó deleción de BRCA1 en ninguno de los tumores TNBC. Sin embargo, se detectó que un 14,5% (7/48) de los tumores presentaron ganancia de una gran región del cromosoma 17q, que involucra el gen BRCA1, además de HER2 (tabla 4). Sin embargo, estos tumores no expresan HER2, sugiriendo que el aumento en el número de copias, obtenido para el gen HER2, no fue suficiente para mostrar expresión de la proteína respectiva. Todos estos tumores fueron TNBC/tipo Basal y 4 de ellos expresaron la proteína BRCA1 con moderada/fuerte intensidad. Probablemente, la fuerza del contexto de las sondas aledañas al gen BRCA1 o la existencia de una microdeleción del gen BRCA1 no pudo ser detectada por algoritmo ADM-1, cuyo promedio del intervalo con aberración es sensible al contexto de las sondas. Por esto, se graficó la razón de log2 (tumor/referencia) de las cuatro sondas que reconoce el gen BRCA1, de cada tumor, excluyendo los tumores con ganancia de una gran región genómica del brazo q del cromosoma 17 (figura 12). 90
114 De esta forma, se supuso que un tumor puede tener deleción del gen BRCA1, si presenta dos o más sondas consecutivas, cercanas a la razón de log2 (tumor/referencia) -1 (figura 12). Este análisis reveló que un 36,5% (15/41) de los tumores TNBC tiene pérdida de al menos un alelo del gen BRCA1 (tabla 5). De estos tumores, un 86,6% (13/15) fueron definidos TNBC/tipo Basal y 2 casos resultaron TNBC/tipo no-basal. Así, un 35% (13/37) de los tumores clasificados como tipo Basal presentan deleción del gen BRCA1. Por otra parte, se encontró que un 80% (12/15) de los tumores con deleción del gen BRCA1, principalmente definidos en el subgrupos TNBC/tipo Basal, presentan una reducida expresión de la proteína BRCA1 en el núcleo. Este hallazgo sugiere que sólo un alelo expresa la proteína BRCA1, ya que el segundo alelo se encuentra delecionado. En cambio, en un 20% (3/15) de los casos con deleción del gen BRCA1 se encontró expresión de la proteína BRCA1 con intensidades moderada/fuerte. Posiblemente, en estos tumores el alelo normal de BRCA1 puede estar expresando isoformas de BRCA1 y no la proteína completa de BRCA1. O bien pueden portar una mutación somática en el gen BRCA1, la cual se expresa en el tumor. Además, 2 de los 3 tumores de portadoras de mutación en el gen BRCA1 en la línea germinal, presentaron deleción del gen BRCA1. En estos dos tumores se identificó una mutación hacia el C-terminal de la proteína BRCA1, sugiriendo que se expresa el alelo mutado no funcional ya que el segundo alelo se encuentra delecionado. Estos resultados indican que una microdeleción, más que una gran deleción de la región genómica que contiene el gen BRCA1, puede revelar la pérdida de heterozigocidad de BRCA1. 91
115 Tabla 4: Tumores de cáncer de mama triple negativo con ganancia de la región cromosómica 17q21.31 que contiene el gen BRCA1. Posición en cromosoma 17 ID Tumor Citobanda Inicio Término Tamaño Locus Clasificación Clasificación según la expresión y (Mb) HER2 subgrupo localización de BRCA1 T_TN48 17q q Ganancia tipo Basal T_TN28 17q q Ganancia tipo Basal Expresión nuclear de BRCA1 T_M156 17q12-17q Ganancia tipo Basal T_TN49 17q12-17q Ganancia tipo Basal T_P4 17q12-17q Ganancia tipo Basal Pérdida de expresión nuclear de BRCA1 T_TN04 17q q Ganancia tipo Basal T43 17q q Neutra tipo Basal Expresión de BRCA1 citoplasmática con/sin expresión nuclear Locus HER2, localizado en la región cromosómica 17q12. HER2 se refiere al receptor 2 del factor de creciemiento epidermal. 92
116 Log 2 (DNA tumor/dna referencia) Sondas de BRCA1 Figura 12: Gráfico de deleción de las 4 sondas dirigidas al gen BRCA1, en la plataforma de microarreglo de oligonucleótido de DNA. En el eje X se indica la razón log2 del DNA tumor versus el DNA de referencia. En el eje Y se muestran las barras de cada sonda para cada tumor. Cada sonda que reconoce el gen BRCA1 se identifica con un color. Sonda 1 (color azul), Sonda 2 (color rojo), Sonda 3 (color verde), Sonda 4 (color morado). 93
117 Tabla 5: Tumores de cáncer de mama triple negativo con deleción de al menos 2 sondas consecutivas del gen BRCA1, en relación a la expresión y localización de la proteína BRCA1. Clasificación según la expresión y localización de BRCA1 TNBC/tipo Basal TNBC/tipo no-basal Expresión nuclear de BRCA1 T_TN31 - Pérdida de expresión nuclear de BRCA1 Expresión de BRCA1 citoplasmática con/sin expresión nuclear T_TN20, T_TN16, T_TN06_2, T_TN17, T41, T_TN32, T24 T_TN38, T_TN39, T_TN34, T_TN07, T11 T_TN46 T_TN19 94
118 4.3.4 Análisis del perfil de frecuencia de los cambios en el número de copias del DNA en los tumores TNBC. A continuación, en la figura 13A, se muestra un gráfico del perfil de cambios en el número de copias del DNA detectadas en los tumores TNBC. En esta figura, se muestran las sondas con ganancia (color rojo) y las sondas con deleción (color verde) a través de todos los cromosomas, indicando el número de tumores que los portan (eje X). El espectro de las sondas de ganancia y de deleción en los tumores TNBC, organizadas según la posición nucleotídica de telómero a telómero, de cada cromosoma, revela regiones cromosómicas con cambios en su número de copias, recurrentes y focalizados entre los tumores TNBC. Además, se identificó un mayor número de regiones genómica de ganancia que de deleción, lo cual sugiere que el genoma de los tumores TNBC puede estar en un estado de ganancia de función, involucrando potenciales oncogenes en la progresión de estos tumores. El perfil de cambios en el número de copias del DNA obtenido de los tumores TNBC difiere del perfil de regiones genómicas alteradas del análisis del genoma de tumores de cáncer de mama hereditario que se muestra en la figura 13B (Carolina Álvarez, manuscrito en preparación). Los tumores de cáncer de mama hereditario, seleccionados por la historia familiar, independiente del inmunofenotipo, presentan rearreglos genómicos menos frecuentes entre ellos a lo largo de todos los cromosomas con similar frecuencia de rearreglos genómicos de ganancia y de deleción a diferencia de los tumores TNBC. Esto sugiere que existen alteraciones genómicas específicas, comunes entre los tumores TNBC, que se diluyen en los análisis de cambios en el número de copias del DNA al evaluar tumores de mama con diferente inmunofenotipo. Los rearreglos genómicos de los tumores TNBC muestra regiones genómicas con límite en el telómero, pero más frecuentes fueron aquellas con límites internos intra-cromosomas. La 95
119 localización del rearreglo genómico en el cromosoma puede relacionarse al mecanismo que produce estos cambios en el número de copias del DNA (Zack y cols., 2013). Otra característica de los rearreglos genómicos (o intervalos genómicos) identificados en los tumores TNBC fue la longitud. Está fue variable entre los tumores TNBC, observándose diferentes límites o puntos de quiebre para una misma región cromosómica. Principalmente, los rearreglos genómicos de ganancia fueron de longitud variable y de gran tamaño en comparación a las regiones genómicas con deleción. La diferencia en la longitud de los rearreglos genómicos de una misma región cromosómica es frecuente de observar, al estudiar el genoma del cáncer, ya que existen distintos sitios susceptibles de quiebre dentro del cromosoma, generándose alteraciones de diferentes longitudes para una misma región del cromosoma, pero involucrando una región mínima de ganancia o de deleción que se comparte entre las muestras que las portan. 96
 Perfil de frecuencia en los tumores TNBC. B) Perfil de frecuencia en tumores de cáncer de mama hereditario.")
. En el eje X se muestran los cromosomas, separados por líneas verticales de color negro.")
120 N de tumores TNBC A) B) X Posición en cromosomas Figura 13: Perfil de frecuencia de los cambios en el número de copias del DNA. A) Perfil de frecuencia en los tumores TNBC. B) Perfil de frecuencia en tumores de cáncer de mama hereditario. En el eje Y se indica el número de tumores TNBC, en la parte superior del gráfico de indican los cambios en el número de copias de ganancia (color rojo) y en la parte inferior se indican las alteraciones genómicas con deleción (color verde). En el eje X se muestran los cromosomas, separados por líneas verticales de color negro. El centrómero es indicado con una línea de color gris, según corresponde. El perfil de frecuencia de cambios en el número de copias del DNA en los tumores de cáncer de mama hereditario pertenece a Carolina Álvarez (manuscrito en preparación). 97
121 Por esto, se caracterizaron los rearreglos genómicos mínimos compartidos entre los tumores TNBC de una misma región cromosómica (Andre y cols., 2009). Así el tumor o los tumores con el rearreglo genómico de menor longitud para una misma región cromosómica fueron determinados como la región mínima compartida. La mayoría de los tumores TNBC mostraron regiones genómicas de ganancia recurrentes en los cromosomas 1p, 2q, 7q, 11p, 11q, 12q, 16p, 17p, 17q, 19p, 19q y 22q. En cambio, los rearreglos genómicos delecionados fueron localizados en los brazos cromosómicos 1q, 6p, 9q, 11q y 19q. Por otra parte, en los tumores TNBC se identificaron 61 rearreglos genómicos en promedio (rango 9-120, en tumores individuales). El promedio de los rearreglos genómicos de ganancia fue de 50 (rango 5-98, en tumores individuales), en tanto, que el promedio para las regiones con deleción fue de 11 por tumor (rango 4-23, en tumores individuales). En la figura 14 se muestra el número de regiones con ganancia y deleción obtenidas por tumor, observándose que sólo el tumor T_TN12 presentó similar número de rearreglos genómicos en ambos estados (ganancia y deleción). Los resultados indican que existe variabilidad en los tumores TNBC, en cuanto al número de aberraciones cromosomales que ellos portan, sugiriendo una alta inestabilidad genómica. 98
122 T_TN10 T_TN15 T_TN48 T_TN31 T_TN02 T_TN03 T_TN44 T_M156 T_TN51 T_TN28 T_TN04 T_TN47 T_TN16 T_TN45 T_P4 T_TN49 T_TN17 T41 T_TN27 T_TN06_2 T_TN46 T42 T_TN32 T_TN43 T_TN20 T_TN08 T_TN38 T24 T_TN06_1 T_TN11 T_TN07 T_TN01 T_TN34 T_TN05 T_TN39 T_TN21_1 T_TN24 T_TN12 T_TN14 T_TN50 T39 T_TN21_2 T_TN19 T43 T_TN42 T5 T_TN09 T11 N de intervalos genómicos 140 N de Intervalos genómicos N de Intervalos con Ganancia N de Intervalos con Deleción Figura 14: Número de intervalos genómicos identificados en cada tumor TNBC. En el gráfico, las barras celestes representan el número total de intervalos genómicos con cambio en su número de copias por tumor TNBC. El número de intervalos en estado de ganancia están indicados con las barras de color rojo. Por su parte, el número de intervalos en estado de deleción se indican con las barras de color verde. 99
123 4.3.5 Identificación de los rearreglos genómicos de ganancia y de deleción recurrentes en los tumores TNBC. En total se identificaron 403 rearreglos genómicos de ganancia, compartidos por un mínimo de 2 tumores TNBC (rango 4,2-81,3% de los tumores TNBC). De estos, 122 (30%) rearreglos genómicos presentaron un tamaño 1 Mb, y las 281 (70%) regiones restantes resultaron <1 Mb. Las regiones genómicas de ganancia portan un número variable de genes, según el cromosoma (relacionado con la densidad de genes que contiene el cromosoma respectivo), y el número de sondas que reconocen un mismo gen. La mediana del porcentaje de rearreglos genómicos de ganancia fue de 20,8% (10/48) en los tumores TNBC. Superior a este valor, se obtuvieron 145 regiones cromosómicas con cambios en su número de copias, distribuidas en todos los cromosomas, que se indican en la figura 15. Por otra parte, en un porcentaje mayor al 50% de los tumores TNBC se encontraron 63 de los 145 rearreglos genómicos con ganancia (figura 15). De estos, dos regiones cromosómicas ubicadas en la citobanda 16p13.3 resultaron las más recurrentes, en un 81,3% de los casos. Por otra parte, distintos rearreglos genómicos con ganancia recurrentes en los tumores TNBC fueron significativamente asociados al subgrupo tipo Basal o a la expresión y localización de BRCA1 que corresponden a: 2q37.3 (20 genes), 4p16.3 (10 genes), 4p16.3-p16.2 (34 genes), 4p16.1 (3 genes), 5q15.33 (15 genes), 7p22.3-p22.2 (19 genes), 8q24.3 (59 genes), 9q34.11-q34.13 (74 genes), 9q34.2-q34.3 (64 genes), 11p15.5 (47 genes), 11p15.4 (5 genes), 11q13.1 (98 genes), 11q13.1-q13.2 (98 genes), 11q13.2-q13.3 (24 genes), 14q32.33 (7 genes), 16p13.3 (148 genes), 16q24.3 (20 genes), 17q25.1 (64 genes), 17q25.3 (60 genes), 19p13.3 (165 genes), 19p13.2 (25 genes), 19p13.11 (3 genes), 20q13.33 (46 genes), 21q22.3 (29 genes), 22q11.21-q11.23 (86 genes), 22q12.2 (8 genes). 100
124 101 Continúa en la siguiente página Ganancia Inicio Término Tamaño (Mb) N de genes/región %TNBC Total % TNBC/tipo basal % TNBC/tipo nobasal T_TN10 T_TN48 T_TN31 T_TN02 T_TN44 T_TM156 T_TN51 T_TN28 T_TN15 T_TN03 T_TN04 T_TN45 T_TN47 T_TN16 T_TN17 T_P4 T_TN49 T41 T_TN27 T_TN06_2 T42 T_TN32 T_TN43 T_TN20 T_TN46 T_TN08 T_TN38 T24 T_TN11 T_TN07 T_TN01 T_TN34 T_TN39 T_TN21_1 T_TN24 T_TN12 T_TN50 T39 T43 T11 T_TN14 T_TN05 T_TN06_1 T_TN21_2 T_TN19 T_TN42 T5 T_TN09 % BRCA1 Expresion nuclear % Ausente/reducido BRCA1 % Expresion citoplasmática BRCA1 1p36.33-p X X X 0 X X X X X X X X X 0 0 X X X X X 0 X 0 0 X 0 X X X X 0 X X p36.33-p X X X 0 0 X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X X 0 X p X X X 0 0 X X X X X X X X 0 0 X X X X X 0 X 0 0 X 0 0 X X X 0 X p X X X X 0 X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X 0 X p X X X 0 X X X X X X X X X X X X X X X 0 0 X 0 0 X X X 0 X p X X X X 0 X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X 0 X p X X X 0 X X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X 0 X p X X X 0 0 X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X 0 X X p X X X 0 X X X X X X X X X X X X X 0 X 0 0 X 0 0 X X X 0 X p X X X 0 0 X X X X X X X X X X X 0 X X X X 0 X p X X X 0 0 X X X X X X X X X X X X X X X p X X X 0 0 X X X X X X X X X X X 0 X 0 0 X 0 0 X X X p X 0 X 0 0 X 0 X X X 0 X X X 0 X X X X p X 0 0 X 0 X 0 X X X 0 X X X X X p X 0 X X 0 X X 0 X X X 0 0 X X X q X X X 0 X X X X X X X 0 X 0 0 X X X 0 X 0 X 0 0 X 0 X X X q X X X 0 X X 0 X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X X X p X 0 0 X 0 0 X X X X 0 0 X X X X p X X X 0 X X 0 X 0 0 X X X 0 0 X 0 0 X 0 0 X X X X q X X X 0 0 X X 0 X X X X X X q X X X 0 X X X X X 0 X X X X 0 X 0 X X 0 X X X 0 X X X q X X X 0 X X X X X 0 X X X X 0 X 0 X X 0 X X X X 0 X X X q X X X 0 X X X X X X X X X 0 0 X X 0 X X X X 0 0 X 0 X X X X X X X X X 0 X p X X X X 0 0 X X 0 X X X p X X X 0 X X 0 X X X X X X 0 0 X X X 0 X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X X X p21.31-p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X 0 X X q X X X 0 X X X X X X X X X X X X 0 0 X 0 0 X 0 0 X X X 0 X X X q X X X 0 0 X X X X X X X X 0 0 X X 0 0 X X X 0 0 X 0 X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X p16.3-p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X 0 X 0 X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 X X 0 0 X X X 0 X 0 X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 X X X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 0 X 0 X X 0 X X X X X X 0 0 X 0 X X X X 0 0 X X 0 0 X q35.2-q X X X 0 X X 0 X 0 0 X X X X 0 X X 0 X p X X 0 X X X 0 X X 0 0 X X X 0 0 X X X p22.3-p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X p X 0 0 X 0 X X 0 X 0 X X X X 0 X 0 X X X X X X p22.2-p X X 0 X X X X 0 X 0 X X X X X 0 0 X 0 0 X X X X X X p X 0 X 0 X X X X X X X X X 0 0 X X 0 X X X X 0 0 X 0 X X X X X 0 0 X 0 X X 0 0 X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 0 X X X 0 X X X X q X X X 0 0 X X X X 0 X X X X X X 0 X X 0 0 X 0 0 X X X X X q21.3-q X X 0 0 X X X X X X 0 X 0 0 X X X X X 0 X q X X 0 0 X X X X X 0 X 0 0 X X X q X X X 0 X X X X X 0 X X X X X X 0 X X 0 0 X 0 X X X 0 X X q X X X 0 X 0 X X X X X X X X 0 X X X X 0 0 X 0 0 X X X X X X X q X 0 X 0 X X 0 X X X X X 0 X X X X X 0 X q X X X X 0 X X 0 0 X X X X p X X X 0 X X X X X X 0 X X X X X X X X X X X 0 0 X X X 0 X p X X X 0 X X X X X X 0 X X X X X X X X X X X X X 0 X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X q X X 0 0 X X 0 0 X X 0 0 X X X X X X X q34.11-q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q34.2-q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X 0 X X 0 X 0 0 X 0 X X X X q X X X 0 X X X X X 0 X 0 0 X X q X X X 0 X X 0 0 X X X 0 X 0 0 X X X X X X X X X X X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X X X 0 X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X 0 0 X X X 0 X X X 0 X X X 0 0 X 0 X X X 0 0 X X X X X X X p X X 0 X X 0 X X X X X X X X p X X 0 0 X 0 X 0 0 X X X X X X p X X 0 0 X 0 X 0 0 X X X X X X 0 X 0 X q12.2-q X X X 0 X X X X X X X X X X X X X X X 0 0 X 0 X X X X X X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X q13.1-q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X 0 X 0 X X X X X q13.2-q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X q X X 0 0 X X X X X 0 0 X X X q X X X X X X X X X 0 0 X 0 X 0 0 X 0 X X X q X X X X X X X X X X 0 X X 0 X X X X 0 X 0 X 0 0 X 0 0 X X X X X X q X X X X X X X X 0 X X X 0 X q X X X X X X X X 0 X X 0 X q X 0 X 0 X X X X X X X 0 0 X X X q X 0 X 0 0 X X X X X X 0 0 X X X X 0 X q X 0 X 0 0 X X X X X X 0 0 X 0 0 X X X q X X X 0 X X X X X X X X X X 0 X X 0 X 0 0 X 0 X X X 0 0 X X q X 0 X 0 X X X X X X X X 0 0 X X X q X X X X 0 0 X X X X X X q X X X 0 X X 0 X X X X X X 0 0 X X X X X 0 X 0 0 X 0 X X X X 0 X X q X 0 X 0 X X X X X X X 0 X 0 0 X 0 X 0 X X X 0 X X X X q X 0 X 0 X X 0 X X X X 0 X 0 0 X 0 X 0 X X X 0 X X X X q X X X 0 X X X X 0 X 0 0 X X 0 0 X X X X q X X X 0 X X X X X X X X X 0 0 X 0 X 0 X X X 0 X X X X X X X X 0 X X q X X X 0 X X 0 X X X X X X 0 0 X 0 X 0 X X X 0 X X X X X X X X 0 X X q X X X 0 X X 0 X X X X X X 0 0 X 0 X 0 X X X 0 X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X 0 X 0 X X X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X Tumores con expresión nuclear de BRCA1 Tumores con ausente/reducida expresión nuclear de BRCA1 Tumores con expresión citoplasmática de BRCA1
125 Tumores con expresión nuclear de BRCA1 Tumores con ausente/reducida expresión nuclear de BRCA1 Tumores con expresión citoplasmática de BRCA1 Ganancia Inicio Término Tamaño (Mb) N de genes/región %TNBC Total % TNBC/tipo basal % TNBC/tipo nobasal T_TN10 T_TN48 T_TN31 T_TN02 T_TN44 T_TM156 T_TN51 T_TN28 T_TN15 T_TN03 T_TN04 T_TN45 T_TN47 T_TN16 T_TN17 T_P4 T_TN49 T41 T_TN27 T_TN06_2 T42 T_TN32 T_TN43 T_TN20 T_TN46 T_TN08 T_TN38 T24 T_TN11 T_TN07 T_TN01 T_TN34 T_TN39 T_TN21_1 T_TN24 T_TN12 T_TN50 T39 T43 T11 T_TN14 T_TN05 T_TN06_1 T_TN21_2 T_TN19 T_TN42 T5 T_TN09 % BRCA1 Expresion nuclear % Ausente/reducido BRCA1 % Expresion citoplasmática BRCA1 14q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X X X X 0 0 X X X X X q X X X 0 X X X X X X 0 0 X 0 X X X X X q X 0 0 X X 0 X 0 0 X X X X X X X X 0 X X X q24.1-q X X X 0 X X X 0 0 X X X X 0 0 X X 0 0 X 0 X 0 0 X 0 0 X X X X 0 X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X p X X X X X X X X X X X X X X 0 X X X X X X X 0 X X 0 X X X X 0 X X X X X X 0 0 X X X 0 X X p X X X 0 X X X X X X X X X X 0 X X X X X X X 0 X X 0 X X X X 0 X X X X X X X 0 X X X 0 X X p X X X 0 X X X X 0 0 X X X 0 0 X X 0 X X X X 0 0 X 0 0 X 0 X 0 X X X X X X X X p X X X 0 X X X X 0 0 X X X 0 0 X X 0 X X X X 0 0 X 0 0 X X X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X 0 X X X X X 0 X X X X X X X X X 0 0 X 0 0 X X X X 0 X p X X 0 X X X X 0 0 X X X X X 0 X X 0 0 X 0 0 X X X 0 X q X X X 0 X X X X X X X X X 0 0 X 0 0 X X 0 X 0 0 X 0 X X X X q13-q X X X 0 X X X X X X X X X 0 0 X X 0 X X X 0 X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X 0 X 0 0 X 0 X X X X X 0 X X X q X X X 0 0 X X X X X 0 X X q23.3-q X X 0 0 X 0 0 X X X X X X X X X X 0 X 0 0 X 0 X X X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X p X X X 0 0 X X X X X X 0 0 X X 0 X X X p X 0 0 X X X 0 0 X X X X X X X p X 0 0 X X X 0 0 X X X X X X X q X X 0 X 0 0 X X X X X 0 0 X 0 0 X q X X 0 X X X X X X 0 X X q X X X 0 X X 0 X X 0 X X X 0 0 X X 0 X X 0 X X X X X q X X 0 0 X X X 0 0 X X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X 0 X X q X X X X X X 0 X X X X X X X X 0 X X X q X X X X X X 0 X X X X 0 0 X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X X 0 X X X X X X X X X X 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X p X X X 0 X X X X X X X X X X 0 X X X X X X X 0 0 X 0 X X X X X X X X 0 0 X X X p X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X X X q13.11-q X X 0 X X X X 0 0 X X X X 0 X 0 X X 0 0 X 0 0 X X X 0 X X q X X 0 X X X X 0 X X X X X 0 X 0 X X 0 0 X 0 0 X X X 0 X X q X X 0 X X X X 0 0 X X X X 0 X 0 X X 0 0 X 0 0 X X X X 0 X X q X X 0 X X X X 0 0 X X X X 0 X 0 X X 0 0 X 0 0 X X X 0 X X X q X X 0 X X X X 0 0 X X X X 0 X X X X p X X X X X X X X X X p X X X 0 X 0 0 X X X X X X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X X 0 X X q11.21-q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X X X X 0 X q12.1-q X X X 0 X X X X X X X 0 X 0 0 X X 0 X X X X 0 0 X 0 X X X 0 X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X X 0 X X q12.2-q X X X 0 X X X X X X 0 X X 0 0 X X X X X X X 0 X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X 0 0 X X q X X X 0 X X X X X X X X X 0 0 X 0 X 0 X X X 0 0 X 0 X X X X X 0 X X q X X X 0 X X X X X X X X X 0 0 X X X X X X X 0 0 X 0 X X X X X 0 X X q13.31-q X X X 0 X X X X X X X X X X 0 X X X X X X X 0 0 X 0 X X X X X 0 X X Figura 15: Regiones genómicas en estado de ganancia en los tumores TNBC. Se indican las regiones cromosómicas, el inicio, término y tamaño (Mb) de los intervalos de ganancia. Además, se encuentran los porcentajes de cada una de estas ganancias en los TNBC, TNBC/tipo basal y TNBC/tipo no-basal (localizados al lado izquierdo). Los porcentajes de estas alteraciones en los tumores agrupados por la expresión y localización de la proteína BRCA1 se encuentran al lado derecho. Las X de color rojo indican que el tumor porta la ganancia. Las celdas blancas indican ausencia de la alteración en el tumores respectivo. En la parte superior de esta figura se encuentran los tumores TNBC. 102
126 Los tamaños de estas regiones cromosómicas fluctuaron entre 0, Mb, con límites principalmente intra-cromosómicos, posiblemente revelando un mecanismo diferente a la disfunción de los telómeros. Estos resultados indican que los tumores TNBC presentan rearreglos genómicos de ganancia recurrentes, y que algunos de estos cambios en el número de copias del DNA están asociados al fenotipo tipo Basal o a la expresión y/o localización de la proteína BRCA1, como se detalla más adelante. Por otra parte, en los tumores TNBC se identificaron 180 rearreglos genómicos con deleción. De las cuales, 97 (53,8%) regiones genómicas fueron encontradas en al menos 2 tumores TNBC (4,2% de los casos) (figura 16). La mediana del porcentaje de los rearreglos genómicos de deleción fue de 6,25% (3/48) en los tumores TNBC, inferior al obtenido para los rearreglos genómicos de ganancia. Este resultado indica que los rearreglos genómicos con deleción fueron menos recurrentes entre los tumores TNBC analizados. Por otra parte, los tamaños de los intervalos genómicos delecionados, generalmente presentaron limites similares entre los tumores o se distanciaban por 2 sondas, a diferencia de los intervalos genómicos de ganancia, cuyos límites fueron variables entre los tumores TNBC. La longitud de los rearreglos genómicos con deleción fluctuó entre Mb, encontrándose con una longitud 1Mb tan sólo 12 regiones genómicas con alteración. Así, regiones genómicas delecionadas con tamaños <1 Mb fueron más redundantes en los tumores de este estudio. Los rearreglos genómicos delecionados en un porcentaje mayor al 20% de los tumores TNBC, corresponden a: 1q21.2 (5 genes), 1q21.3 (4 genes), 6p22.1-p21.33 (17 genes), 6p11.2 (1 genes), 9q33.1 (2 genes), 9q33.1 (2 genes), 9q33.1 (1 genes), 11q22.3 (3 genes), 17q21.32 (3 genes), 19q13.3 (9 genes), 19q13.3 (3 genes), 19q13.41 (1 genes), 19q13.41 (5 genes), 19q13.41 (5 genes). 103
127 104 Figura 16: Regiones genómicas en estado de deleción en los tumores TNBC. Se indican las regiones cromosómicas, el inicio, término y tamaño (Mb) de los intervalos de deleción. Además, se encuentran los porcentajes de cada una de estas deleciones en los TNBC, TNBC/tipo Basal y TNBC/tipo no-basal (localizados al lado izquierdo). Los porcentajes de estas alteraciones en los tumores agrupados por la expresión y localización de la proteína BRCA1 se encuentran al lado derecho. Las X de color verde indican que el tumor porta la deleción. Las celdas blancas indican ausencia de la alteración en el tumores respectivo. En la parte superior de esta figura se encuentran los tumores TNBC. T_TN10 T_TN48 T_TN31 T_TN02 T_TN44 T_TM156 T_TN51 T_TN28 T_TN15 T_TN03 T_TN04 T_TN45 T_TN47 T_TN16 T_TN17 T_P4 T_TN49 T41 T_TN27 T_TN06_2 T42 T_TN32 T_TN43 T_TN20 T_TN46 T_TN08 T_TN38 T24 T_TN11 T_TN07 T_TN01 T_TN34 T_TN39 T_TN21_1 T_TN24 T_TN12 T_TN50 T39 T43 T11 T_TN14 T_TN05 T_TN06_1 T_TN21_2 T_TN19 T_TN42 T5 T_TN09 1p X X p X X p X X p X X p X 0 0 X X X q X X X X X X 0 X X X X 0 X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X q X X 0 0 X X 0 X X 0 X X X X X X X q X X 0 0 X 0 0 X X X 0 X X X 0 X X X X X X 0 X q X X X X X X q31.1-q X X q X 0 X X X p X 0 0 X q36.3-q X X q X X p X X X p X 0 0 X X X p X 0 0 X X X X X p X X X X X p X X X X p X X p X X p X X q X X q X X p22.1-p X 0 0 X X X X X X 0 X 0 0 X X X X p X 0 X X X X X X X X X X X X X X 0 X 0 0 X 0 X X 0 0 X 0 X X X X 0 X X p X X q X X q X X X q X X X X 0 X X X q X X q X X X X X X 0 X X X X X 0 0 X 0 X X X q X X 0 0 X X 0 X X X X X 0 0 X 0 X X X q X 0 0 X X X X X X 0 0 X 0 X X X q X X 0 0 X X 0 X X X q X X X X 0 X q X X X X p X X q X X X X X q X X X q X X X X 0 0 X X X X q X X X 0 0 X X X X X X X X 0 0 X 0 0 X X 0 X 0 0 X 0 X X X X X q X X X q X X p X X q X X X X 0 0 X X 0 0 X q X 0 0 X q X X 0 X 0 X q X X X q X X q X X X X 0 0 X 0 0 X X q X 0 0 X X q X X X p X X X p X X p X X X p X X q X X X q X X 0 X X X X X X q X X X 0 X p X X X q X X q X 0 0 X X X q X X q X X X 0 X 0 0 X X 0 X X X X q X X X q X X X q X X X X q X X X X X X X X q X X X X p X X p13.3-p X X X X 0 0 X X X X p X X X p X X X X 0 X 0 0 X p X X X p X X X X X q X X X X q X 0 0 X X X 0 X X X X q X 0 X 0 X 0 X 0 0 X X q X X X q X X q X X 0 X X X 0 X X X X X 0 0 X 0 0 X X 0 X 0 0 X X X X q X 0 0 X X X 0 0 X X 0 X 0 0 X X X q X 0 X X X X X X X 0 X 0 0 X X X 0 X X X X X 0 X X X X X 0 0 X X q X 0 0 X X X X X X X 0 X 0 0 X X X q X X X X 0 X X 0 X 0 0 X X X q X X X q X X X X q X X q X X X q X 0 X q X X X X X X q X X q X 0 0 X X X X q X X X X q X X X q X 0 X % Ausente/reducido BRCA1 % Expresión citoplasmática BRCA1 Tamaño (Mb) Término Inicio Deleción % BRCA1 Expresión nuclear Tumores con expresión nuclear de BRCA1 Tumores con ausente/reducida expresión nuclear de BRCA1 Tumores con expresión citoplasmática de BRCA1 N de genes/región %TNBC Total % TNBC/tipo basal % TNBC/tipo no-basal
128 Alguno de los cuales fueron asociados al subtipo tipo Basal o a la expresión y localización de BRCA1 (se detalla más adelante). Además, del rearreglo genómico con deleción más recurrente que fue 1q21.1 (91,7%) entre los tumores TNBC (figura 16). Esta región genómica por primera vez es descrita como recurrente entre los tumores TNBC. La alta recurrencia de algunos rearreglos genómicos de ganancia y de deleción en los tumores TNBC, sugiere que estos tumores de mama presentan un patrón de cambios en el número de copias del DNA particular, que podrían permitir diferenciarlo de otros tumores de cáncer de mama. Aún más, estas regiones genómicas alteradas, frecuentes en los tumores TNBC, pueden contener genes, ya sea protoncogenes o genes supresores de tumor, funcionalmente importantes para la progresión de este tipo de tumor Cambios en el número de copias del DNA diferencia los tumores TNBC/tipo Basal de los TNBC/tipo no-basal. Una vez determinados los rearreglos genómicos de ganancia y deleción en los tumores TNBC, se identificaron los cambios en el número de copias del DNA que diferencian los TNBC/tipo Basal de los TNBC/tipo no-basal. Los tumores TNBC/tipo Basal en promedio concentraron 65,5 rearreglos genómicos versus 44,2 obtenidos para los tumores TNBC/tipo no- Basal. Esta diferencia, en el promedio de rearreglos genómicos, se mantiene al considerar el promedio de los rearreglos genómicos de ganancia ( TNBC/tipo Basal : promedio 53,8 versus TNBC/tipo no-basal : promedio 33,7). En cambio, el promedio de los rearreglos genómicos con deleción fue similar entre ambos grupos de tumores ( TNBC/tipo Basal : promedio 11,6 versus TNBC/tipo no-basal : promedio 10,5). Esto demuestra que los tumores TNBC/tipo Basal presentan un mayor número de rearreglos genómicos, principalmente con ganancia en el número 105
129 de copias del DNA, sugiriendo que ellos tienen mayor inestabilidad genómica en comparación con los tumores TNBC/tipo no-basal. Los rearreglos genómicos con alteración en su número de copias, específicos de cada grupo de tumor TNBC, definido por la expresión de las citoqueratinas tipo Basal y EGFR fue determinada mediante la prueba estadística del test de Fisher con un valor de p <0.05. En total, 12 regiones genómicas con ganancia difieren significativamente entre los dos grupos de tumor TNBC. Los rearreglos genómicos de ganancia asociados a los tumores TNBC/tipo Basal corresponde a: 5q35.2-q35.3 (14 genes), 7q21.3-q22.1 (5 genes), 7q22.1 (10 genes), 7q22.1 (36 genes), 8p21.3 (18 genes), 10q24.33 (3 genes), 15q15.1 (43 genes), 15q15.1 (7 genes), 16p13.3 (26 genes), 16p11.2 (67 genes), 16p11.2 (3 genes), y 19q13.43 (7 genes). Estas regiones genómicas con alteración fueron encontradas en un rango porcentual de 30-65% en los tumores TNBC/tipo Basal a diferencia de los TNBC/tipo no-basal (rango porcentual de 0-27%). De estas regiones de ganancia, el rearreglo genómico identificado en el cromosoma 10q, ha sido asociado al subgrupo tipo Basal (Adelaide y cols., 2007). Además, nuevos rearreglos genómicos de ganancia fueron identificados en los TNBC/tipo Basal en este estudio. Por otra parte, dos rearreglos genómicos con deleción fueron asociados significativamente a los tumores TNBC/tipo no-basal. Estas regiones corresponden a: 11q14.3 (2 genes) y 12q13.3 (3 genes). En este estudio no se identificaron rearreglos genómicos de deleción asociados significativamente a los tumores TNBC/tipo Basal. Sin embargo, regiones genómicas con perdida en su número de copias, en una frecuencia mayor al 10%, fueron únicamente detectadas en los casos TNBC/tipo Basal que corresponden a: 3p21.31 (3 genes), 7q22.1 (3 genes), 11q13.1 (4 genes), 19p13.2 (5 genes) y 22q12.3 (2 genes). 106
130 Estos resultados indican que los tumores TNBC/tipo Basal son genómicamente diferentes a los TNBC/tipo no Basal, con cambios en el número de copias del DNA específicos del subgrupo de cáncer de mama triple negativo. Además, sugiere que las diferencias en el patrón de expresión génica de estos dos grupos de tumores de mama pueden estar, en parte, concertado por los diferentes cambios en el número de copias del DNA. Por otra parte, este análisis revela la heterogeneidad genómica de los tumores TNBC Agrupamiento jerárquico no supervisado en base a los genes con ganancia y deleción identificados en los tumores TNBC. Con el fin de determinar el agrupamiento natural que adquieren los tumores TNBC se realizó un agrupamiento jerárquico no supervisado, utilizando los genes con cambio en el número de copias del DNA, con ganancia y deleción. Para esto, se seleccionaron los genes de rearreglos genómicos con ganancia en su número de copias, si el 100% de las sondas dirigidas al gen, se encontraron en el mismo estado. Por otra parte, los genes de regiones genómicas con pérdida en su número de copias fueron escogidos si al menos una de las sondas del gen se encontró en estado de deleción. Además, se excluyeron los genes presentes en menos de 2 tumores. Con este criterio se obtuvo un total de 7293 genes, ya sea en estado de ganancia o deleción, identificados en 3 o más tumores TNBC. De estos, 6749 (92,5%) corresponden a genes localizados en regiones genómicas con ganancia y 544 (7,5%) genes ubicados en rearreglos genómicos con deleción. Esta diferencia fue producto del gran tamaño de los rearreglos genómicos con ganancia y su mayor frecuencia entre los tumores TNBC, con un promedio de 30 genes por región genómica de ganancia comparado con un promedio de 5 genes por región genómica con deleción. El agrupamiento jerárquico no supervisado estratificó los tumores TNBC (n = 48), basándose en los genes con cambios en su número de copias en dos grupos principales. Estos dos 107
131 grupos se diferenciaron principalmente por el número de genes con aberración o alteración (figura 17A). La rama de la izquierda, fue representada por 2200 genes con aberración, indicando que los tumores que conforman este grupo contienen un menor número de genes con cambios en su número de copias (Bajo CNA, CNA: Alteración en el número de copias). En cambio, la rama de la derecha del agrupamiento jerárquico no supervisado, fue representada por 5000 genes con alteración. Indicando que los tumores que conforman esta rama contiene un mayor número de genes con cambios en el número de copias del DNA (Alto CNA). Esto sugiere que el número de rearreglos cromosomales fue el determinante del agrupamiento jerárquico no supervisado. El grupo definido con bajo CNA fue formado por un 43,75% (21/48) de los tumores TNBC y el grupo con alto CNA por un 56,25% (27/48) de los casos (figura 17A). De esta manera, ambas grupos quedaron conformados con proporciones similares de tumores TNBC. Estos resultados sugieren que el grupo de tumores con bajo CNA presentan un genoma menos inestable, en comparación con el grupo de tumores con alto CNA, ya que un menor número de genes se encuentran con cambios en su número de copias. En cambio, el grupo de tumores con alto CNA, portan un genoma con alta inestabilidad. En la rama denominada bajo CNA, se localizaron 9 de los 10 tumores con histología tipo especial, incluyendo los tumores T_TN21_1 y T_TN21_2 que provienen de la misma paciente, indicando que conservan alteraciones genéticas similares. La alta proporción de los carcinomas de tipo especial, agrupados en la rama con bajo CNA, sugiere que conservan un perfil con un bajo número de rearreglos genómicos similares, a diferencia de los carcinomas de tipo no especial de los tumores TNBC. 108
132 Porcentaje de sobrevida total T_TN12 T_TN06_1 T_TN05 T_TN43 T_TN42 T_TN08 T_TN20 T_TN02 T_TN21_2 T_TN17 T_TN14 T_TN01 T_TN07 T_TN21_1 T_TN11 T_TN24 T_TN16 Genes con alteración en su numero de copias T_TN09 T39 T11 T_TN19 T_P4 T_TN31 T_TN28 T_TN49 T_M156 T_TN39 T_TN38 T_TN47 T_TN06_2 T_TN03 T_TN15 T_TN10 T42 T5 T41 T_TN44 T_TN45 T_TN50 T_TN46 T_TN04 T_TN48 T_TN32 T_TN34 T_TN27 T43 T24 T_TN51 Bajo CNA Alto CNA A) B) Percent survival Genes Branch con A Bajo CNA Branch Genes con B Alto CNA Day Hazard ratio Dias Figura 17: A) Agrupamiento jerárquico no supervisado de los tumores TNBC, en base la ganancia y deleción de genes alterados en su número de copias. En la figura se indican las dos principales ramas, denominadas Bajo CNA y Alto CNA. B) Gráfico de la sobrevida total de los casos con Bajo CNA y Alto CNA. 109
133 Por otra parte en relación a los tumores provenientes de portadoras de mutación en alguno de los dos genes de alto riesgo al cáncer de mama, se encontró que el grupo con alto CNA concentra un 86% (6/7) de los tumores con mutación en el gen BRCA1 (n = 2) y el gen BRCA2 (n = 4). Solo el tumor T_TN17, proveniente de una paciente portadora de una mutación en el gen BRCA1 en la línea germinal fue agrupado en la rama con bajo CNA. Esto muestra que los tumores provenientes de pacientes con mutación en el gen BRCA1 o BRCA2 presentan un alto número de aberraciones genómicas, sugiriendo una alta inestabilidad genómica. Posteriormente se evaluó la sobrevida total de las pacientes en relación a los dos grupos obtenidos del agrupamiento jerárquico no supervisado. Este análisis mostró que los tumores de la rama con bajo CNA presentan una mejor sobrevida total, en comparación con los tumores de la rama con alto CNA, que mostraron una menor sobrevida total, desde la fecha del diagnóstico de cáncer de mama (Hazard ratio = 0,3379). Sin embargo estas diferencias no fueron estadísticamente significativas posiblemente por el bajo número de nuestras (figura 17B). Por otra parte, se determinó si en nuestro grupo de genes con alteración en su número de copias se encontraban algunos de los genes conductores del cáncer, descritos por Volgestein y cols., (2013). Estos autores, describieron un total de 65 genes conductores de los cuales 30 corresponden a oncogenes y 25 a genes supresores de tumor. Principalmente estos genes se han encontrado mutados somáticamente en diferentes tipos de cáncer, entre ellos el cáncer de mama (Volgestein y cols., 2013). Nuestro análisis reveló que en ambas ramas con bajo CNA y alto CNA existe ganancia de los genes ABL1 (9q34.12), AKT1 (14q32.33), CBL (11q23.3), CSF1R (5q33.1), FGFR3 (4p16.3), U2AF1 (21q22.3), todos definidos como oncogenes y conductores del cáncer. Se ha descrito que el gen ABL1 codifica una proteína involucrada en el ciclo celular/apoptosis. Los genes AKT1, CBL, CSF1R y FGFR3 codifican para proteínas implicadas 110
134 en la vía de señalización de las PI3K y en la vía de señalización de RAS, en cambio el gen U2AF1 codifica para una proteína que participa en la regulación transcripcional. Estos genes fueron localizados en rearreglos genómicos de ganancia, lo cual sugiere que podrían estar sobreexpresados en los tumores TNBC que los portan. Además, solo en la rama con alto CNA se identificó el gen MPL en una región con ganancia en su número de copias, el cual también ha sido descrito como un gen conductor del cáncer. El agrupamiento jerárquico no supervisado de los tumores TNBC no mostró una clara separación en relación a los subtipos TNBC/tipo Basal y los TNBC/tipo no-basal (p-value = 0,1736) (figura 18). Sin embargo, 7 de los 11 tumores TNBC/tipo no-basal se agruparon en la rama con bajo CNA. De esta forma, el grupo con bajo CNA concentra un 67% (14/21) de los tumores TNBC/tipo Basal y un 33,3% (7/21) fueron TNBC/tipo no-basal. A diferencia del grupo de tumores que conforma la rama con alto CNA, compuesta de un 85,7% (23/27) de los tumores TNBC/tipo Basal y de un 14,8% (4/27) de los casos TNBC/tipo no-basal. Esta tendencia sugiere que los tumores TNBC/tipo Basal son heterogéneos y que diferencias en los cambios en el número de copias del DNA podrían ayudar a homogenizar estos tumores de mama que presentan un perfil de expresión génica similar. Otra diferencia, se observa al evaluar solo la expresión de los marcadores del tipo Basal. Se encontró que un 50% (7/14) de los casos TNBC/tipo Basal con bajo CNA, expresan los tres marcadores CK5+/CK14+/EGFR+ conjuntamente en el mismo tumor, a diferencia de un 17,4% (4/23) de los TNBC/tipo Basal del grupo con alto CNA. Por otra parte, el grupo con bajo CNA presenta mayor intensidad de tinción de EGFR en comparación con los tumores de alto CNA, sugiriendo que estos tumores pueden ser susceptibles a tratamientos con inhibidores de la señalización de EGFR. Es importante 111
135 mencionar que en este grupo de tumores TNBC no se identificó EGFR amplificado, sin embargo, si se encontró expresado en un 43% de los casos TNBC. En relación a la expresión y localización de BRCA1, si se observa una clara separación en el agrupamiento jerárquico no supervisado de los tumores TNBC. En la figura 40, se observó que la rama con bajo CNA, está conformada principalmente por tumores con expresión citoplasmática de BRCA1 con y sin expresión nuclear (76%, 16/21 tumores bajo CNA). En cambio, el grupo con alto CNA concentró los tres grupos de tumores TNBC basados en la expresión y localización de BRCA1. Por lo tanto, el grupo con alto CNA fue constituido por un 90% (9/10) de los tumores con expresión nuclear de BRCA1, 73% (11/15) de los tumores con pérdida de expresión nuclear de BRCA1 y un 30% de los tumores con expresión citoplasmática con y sin expresión nuclear de BRCA1. Esto indica que la rama con alto CNA es heterogénea, en relación a la expresión y localización de BRCA1, pero con menor proporción de tumores con expresión citoplasmática de BRCA1, además de concentrar los tumores TNBC/tipo Basal. 112



136 T_TN12 T_TN06_1 T_TN05 T_TN43 T_TN42 T_TN08 T_TN20 T_TN02 T_TN21_2 T_TN17 T_TN14 T_TN01 T_TN07 T_TN21_1 Genes con alteración en su numero de copias T_TN11 T_TN24 T_TN16 T_TN09 T39 T11 T_TN19 T_P4 T_TN31 T_TN28 T_TN49 T_M156 T_TN39 T_TN38 T_TN47 T_TN06_2 T_TN03 T_TN15 T_TN10 T42 T5 T41 T_TN44 T_TN45 T_TN50 T_TN46 T_TN04 T_TN48 T_TN32 T_TN34 T_TN27 T43 T24 T_TN51 Bajo CNA Alto CNA Tipo Basal tipo no-basal Expresión núcleo/citoplasma Perdida de expresión nuclear Expresión nuclear Figura 18: Agrupamiento jerárquico no supervisado de los tumores TNBC, en relación a la expresión y localización de BRCA1. En la figura se indican las dos principales ramas, denominadas Bajo CNA y Alto CNA. Además, se indica la expresión y localización de BRCA1 en los tumores TNBC. Los rectángulos naranja indican los tumores con expresión citoplasmática de BRCA1 con y sin expresión nuclear. Los rectángulos rojos indican pérdida de expresión de BRCA1 nuclear. Los rectángulos azules indican los casos con expresión nuclear de BRCA1. Además, se muestran los tumores TNBC/tipo Basal (rectángulos verdes) y los tumores TNBC/tipo no-basal (rectángulos celestes). 113
Existe el carcinoma de mama con fenotipo basal?
 Existe el carcinoma de mama con fenotipo basal? David Hardisson Dpt. de Anatomia Patologica c Luminal, ER+ d ERBB2 e Basal f Normal CÁNCER DE MAMA CON FENOTIPO BASAL Definición, espectro morfológico.
Existe el carcinoma de mama con fenotipo basal? David Hardisson Dpt. de Anatomia Patologica c Luminal, ER+ d ERBB2 e Basal f Normal CÁNCER DE MAMA CON FENOTIPO BASAL Definición, espectro morfológico.
SIMPOSIO DE MAMA DIAGNÓSTICO Y SUBTIPOS MOLECULARES
 UNIVERSIDAD DE COSTA RICA HOSPITAL SAN JUAN DE DIOS SIMPOSIO DE MAMA DIAGNÓSTICO Y SUBTIPOS MOLECULARES Dra. Jeannina Álvarez Yannarella Residente de Patología INFORMACIÓN CLÍNICA Sexo y edad Antecedentes
UNIVERSIDAD DE COSTA RICA HOSPITAL SAN JUAN DE DIOS SIMPOSIO DE MAMA DIAGNÓSTICO Y SUBTIPOS MOLECULARES Dra. Jeannina Álvarez Yannarella Residente de Patología INFORMACIÓN CLÍNICA Sexo y edad Antecedentes
CÁNCER DE SENO PRUEBAS GENÉTICAS
 CÁNCER DE SENO PRUEBAS GENÉTICAS La identificación y entendimiento de la genética del cáncer de seno permite individualizar el manejo de la enfermedad así como estudiar a las familias para ofrecer opciones
CÁNCER DE SENO PRUEBAS GENÉTICAS La identificación y entendimiento de la genética del cáncer de seno permite individualizar el manejo de la enfermedad así como estudiar a las familias para ofrecer opciones
INTRODUCCIÓN Linfoma Centroblástico
 INTRODUCCIÓN Los Linfomas No Hodgkin constituyen un grupo heterogéneo de neoplasias malignas linfoproliferativas con una biología y comportamiento clínico diferente. Se produjeron 55,000 casos nuevos de
INTRODUCCIÓN Los Linfomas No Hodgkin constituyen un grupo heterogéneo de neoplasias malignas linfoproliferativas con una biología y comportamiento clínico diferente. Se produjeron 55,000 casos nuevos de
INTRODUCCION A LA BIOLOGIA CELULAR Y MOLECULAR
 INTRODUCCION A LA BIOLOGIA CELULAR Y MOLECULAR - BIOLOGIA CELULAR - Histología Tejidos 1 Tejidos Tejidos 2 Epitelio Clasificación Cilíndrica Cúbica Plana 3 Epitelios simples Plano simple Cúbico simple
INTRODUCCION A LA BIOLOGIA CELULAR Y MOLECULAR - BIOLOGIA CELULAR - Histología Tejidos 1 Tejidos Tejidos 2 Epitelio Clasificación Cilíndrica Cúbica Plana 3 Epitelios simples Plano simple Cúbico simple
CARCINOMAS TRIPLE NEGATIVO (TN) Características histomorfológicas. Dra. Patricia Calafat Servicio de Patología Hospital Privado Córdoba
 Características histomorfológicas. Dra. Patricia Calafat Servicio de Patología Hospital Privado Córdoba") CARCINOMAS TRIPLE NEGATIVO (TN) Características histomorfológicas Dra. Patricia Calafat Servicio de Patología Hospital Privado Córdoba INTRODUCCIÓN Tumores de mama Infiltrantes Grupo heterogéneo de tumores
CARCINOMAS TRIPLE NEGATIVO (TN) Características histomorfológicas Dra. Patricia Calafat Servicio de Patología Hospital Privado Córdoba INTRODUCCIÓN Tumores de mama Infiltrantes Grupo heterogéneo de tumores
 CARCINOMA ADENOIDE QUÍSTICO DEFINICIÓN Tumor maligno epitelial, derivado de glándulas salivares o de glándulas similares de otras localizaciones compuesto por células modificadas mioepiteliales (adluminales)
CARCINOMA ADENOIDE QUÍSTICO DEFINICIÓN Tumor maligno epitelial, derivado de glándulas salivares o de glándulas similares de otras localizaciones compuesto por células modificadas mioepiteliales (adluminales)
Módulo de MAMA. Ronda nº 2
 SEAP Calle Ancora, 3, 2º B 28045 MADRID Tfno. y Fax 91 539 86 28 Mail: seap@seap.es Programa de Garantía de Calidad en Patología Módulo de MAMA Ronda nº 2 Antígeno probado: P63 Tejido probado: Tejido mamario
SEAP Calle Ancora, 3, 2º B 28045 MADRID Tfno. y Fax 91 539 86 28 Mail: seap@seap.es Programa de Garantía de Calidad en Patología Módulo de MAMA Ronda nº 2 Antígeno probado: P63 Tejido probado: Tejido mamario
BUSQUEDA DE MUTACIONES EN LOS GENES BRCA1 Y BRCA2, EN PACIENTES CON CANCER DE MAMA HEREDITARIO
 CONICYT: Repositorio Institucional: Ficha de Iniciativa de CIT (Ciencia, Tecnología e Innovación) 1 FONDECYT-REGULAR - 2001-1011076 BUSQUEDA DE MUTACIONES EN LOS GENES BRCA1 Y BRCA2, EN PACIENTES CON CANCER
CONICYT: Repositorio Institucional: Ficha de Iniciativa de CIT (Ciencia, Tecnología e Innovación) 1 FONDECYT-REGULAR - 2001-1011076 BUSQUEDA DE MUTACIONES EN LOS GENES BRCA1 Y BRCA2, EN PACIENTES CON CANCER
CÁNCER DE MAMA. Modelo de aplicación de la genética y biología molecular. Dr. Abelardo Arias Velásquez
 V SIMPOSIO ANDINO DE LABORATORIO Y MEDICINA CÁNCER DE MAMA Modelo de aplicación de la genética y biología molecular Dr. Abelardo Arias Velásquez Jefe de la Unidad de Genética y Biología Molecular Instituto
V SIMPOSIO ANDINO DE LABORATORIO Y MEDICINA CÁNCER DE MAMA Modelo de aplicación de la genética y biología molecular Dr. Abelardo Arias Velásquez Jefe de la Unidad de Genética y Biología Molecular Instituto
Enrique Lerma Hospital de la Santa Creu i Sant Pau Barcelona. Patología de la Mama en 2016: amenazas vs Oportunidades
 Enrique Lerma Hospital de la Santa Creu i Sant Pau Barcelona Patología de la Mama en 2016: amenazas vs Oportunidades (auto)amenazas vs Oportunidades La Anatomía Patológica es la rama de la Medicina que
Enrique Lerma Hospital de la Santa Creu i Sant Pau Barcelona Patología de la Mama en 2016: amenazas vs Oportunidades (auto)amenazas vs Oportunidades La Anatomía Patológica es la rama de la Medicina que
CICLO CELULAR Y SU REGULACIÓN G1
 G2 M S CICLO CELULAR Y SU REGULACIÓN G1 EXPRESIÓN de GENES Para dividirse, la célula recibe señales, las transmite y las interpreta como EXPRESIÓN de GENES ESPECÍFICOS para poder pasar por cada una de
G2 M S CICLO CELULAR Y SU REGULACIÓN G1 EXPRESIÓN de GENES Para dividirse, la célula recibe señales, las transmite y las interpreta como EXPRESIÓN de GENES ESPECÍFICOS para poder pasar por cada una de
Cáncer. Dr. Monterrubio. Capitulo 14
 Cáncer Capitulo 14 Cáncer 1. Caracterizado por la división celular sin control 2. ~1.5 millón/año de americanos diagnosticados (0.5 millón/año mueren por la enfermedad) 3. 10% de canceres con predisposición
Cáncer Capitulo 14 Cáncer 1. Caracterizado por la división celular sin control 2. ~1.5 millón/año de americanos diagnosticados (0.5 millón/año mueren por la enfermedad) 3. 10% de canceres con predisposición
con cáncer tipo basal podrían requerir terapias sistémicas más agresivas (Gradishar, 2005; Neve et al., 2006).
.") I. INTRODUCCIÓN El cáncer de mama (CaMa) ocupa el primer lugar en causa de muerte por cáncer en mujeres a nivel mundial, con más de 410,000 defunciones cada año (Ferlay et al., 2001). En México y según
I. INTRODUCCIÓN El cáncer de mama (CaMa) ocupa el primer lugar en causa de muerte por cáncer en mujeres a nivel mundial, con más de 410,000 defunciones cada año (Ferlay et al., 2001). En México y según
Es considerada como una de las características funcionales principales de las células.
 DIVISIÓN CELULAR Es considerada como una de las características funcionales principales de las células. El crecimiento y desarrollo adecuados de los organismos vivos depende del crecimiento y multiplicación
DIVISIÓN CELULAR Es considerada como una de las características funcionales principales de las células. El crecimiento y desarrollo adecuados de los organismos vivos depende del crecimiento y multiplicación
Nombre de la asignatura: BIOLOGIA CELULAR Y MOLECULAR DEL CANCER. Miembros del equipo docente: Dr M Monzo Dr A Navarro Dra Carmen Muñoz, Dr.
 Facultat de Medicina PLA DOCENT DE L ASSIGNATURA DATOS GENERALES Nombre de la asignatura: BIOLOGIA CELULAR Y MOLECULAR DEL CANCER Código: 571848 Tipo: Optativa Impartición: Facultad de Medicina UB Departamentos
Facultat de Medicina PLA DOCENT DE L ASSIGNATURA DATOS GENERALES Nombre de la asignatura: BIOLOGIA CELULAR Y MOLECULAR DEL CANCER Código: 571848 Tipo: Optativa Impartición: Facultad de Medicina UB Departamentos
TERAPIA ONCOLÓGICA " UNA VISIÓN DIFERENTE "
 TERAPIA ONCOLÓGICA " UNA VISIÓN DIFERENTE " M. C. Victor Alex Palacios Cabrejos Oncólogo Clínico Servicio de Hematología - Oncología Clínica Hospital Nacional Cayetano Heredia SUMARIO 1. Epidemiología
TERAPIA ONCOLÓGICA " UNA VISIÓN DIFERENTE " M. C. Victor Alex Palacios Cabrejos Oncólogo Clínico Servicio de Hematología - Oncología Clínica Hospital Nacional Cayetano Heredia SUMARIO 1. Epidemiología
GUÍA CLÍNICA PARA PREVENCIÓN Y SEGUIMIENTO DE CÁNCER DE MAMA 2015
 GUÍA CLÍNICA PARA PREVENCIÓN Y SEGUIMIENTO DE CÁNCER DE MAMA 2015 Tomada y adaptada de MSAL Complementaria de Guía de Examen periódico de salud del adulto 2015. EPIDEMIOLOGÍA El cáncer de mama es la primera
GUÍA CLÍNICA PARA PREVENCIÓN Y SEGUIMIENTO DE CÁNCER DE MAMA 2015 Tomada y adaptada de MSAL Complementaria de Guía de Examen periódico de salud del adulto 2015. EPIDEMIOLOGÍA El cáncer de mama es la primera
Informe técnico del proyecto. de diagnóstico genético de pacientes con. síndrome de Dravet y espectros asociados
 Informe técnico del proyecto de diagnóstico genético de pacientes con síndrome de Dravet y espectros asociados Introducción El objetivo principal de este proyecto es realizar un diagnóstico genético a
Informe técnico del proyecto de diagnóstico genético de pacientes con síndrome de Dravet y espectros asociados Introducción El objetivo principal de este proyecto es realizar un diagnóstico genético a
DOCTORADO EN CIENCIAS MEDICAS
 U N I V E R S I D A D D E C O L I M A DOCTORADO EN CIENCIAS MEDICAS ESTUDIO COMPARATIVO ENTRE LA FORSKOLINA VERSUS CROMOGLICATO DE SODIO EN LA PREVENCION DE LAS CRISIS ASMATICAS EN NIÑOS Y ADULTOS. TESIS
U N I V E R S I D A D D E C O L I M A DOCTORADO EN CIENCIAS MEDICAS ESTUDIO COMPARATIVO ENTRE LA FORSKOLINA VERSUS CROMOGLICATO DE SODIO EN LA PREVENCION DE LAS CRISIS ASMATICAS EN NIÑOS Y ADULTOS. TESIS
CÁNCER DE MAMA CÁNCER DE OVARIO
 CÁNCER DE MAMA CÁNCER DE OVARIO Es el tipo de cáncer que afecta a más mujeres en el mundo. Se diagnostican más de 500.000 nuevos casos cada año, con tasas de incidencia en crecimiento. 1 de cada 8 mujeres
CÁNCER DE MAMA CÁNCER DE OVARIO Es el tipo de cáncer que afecta a más mujeres en el mundo. Se diagnostican más de 500.000 nuevos casos cada año, con tasas de incidencia en crecimiento. 1 de cada 8 mujeres
patológico en cáncer de mama
 Actualizaciones en diagnóstico patológico en cáncer de mama Dra. Leonor Moyano Sch Dra. Laura Carreño T Dra. Valeria Cornejo Dr. Arturo Espinoza N Dr. Pablo Matamala Dra. Verónica Sanhueza L Temario 1.
Actualizaciones en diagnóstico patológico en cáncer de mama Dra. Leonor Moyano Sch Dra. Laura Carreño T Dra. Valeria Cornejo Dr. Arturo Espinoza N Dr. Pablo Matamala Dra. Verónica Sanhueza L Temario 1.
Cáncer de mama hereditario. Luis Manuel Delgado Martínez UGC ginecología. Huércal Overa 2013
 Cáncer de mama hereditario Luis Manuel Delgado Martínez UGC ginecología. Huércal Overa 2013 Epidemiología El cáncer de mama (CM) es un problema importante, siendo el más frecuente de los cánceres en mujeres.
Cáncer de mama hereditario Luis Manuel Delgado Martínez UGC ginecología. Huércal Overa 2013 Epidemiología El cáncer de mama (CM) es un problema importante, siendo el más frecuente de los cánceres en mujeres.
HER2 en Cancer Gastrico
 HER2 en Cancer Gastrico Dr. Alejandro Corvalan Laboratorio de Patologia Molecular y Epidemiologia Centro de Investigaciones Medicas Departamento de Hematoloiga &Oncologia P.Universidad Catolica de Chile
HER2 en Cancer Gastrico Dr. Alejandro Corvalan Laboratorio de Patologia Molecular y Epidemiologia Centro de Investigaciones Medicas Departamento de Hematoloiga &Oncologia P.Universidad Catolica de Chile
AGRADECIMIENTOS... VII RESUMEN... XIII RESUM... XV SUMMARY... XVII ABREVIATURAS... XIX ÍNDICE DE CONTENIDO... XXIII ÍNDICE DE FIGURAS...
 ÍNDICE AGRADECIMIENTOS... VII RESUMEN... XIII RESUM... XV SUMMARY... XVII ABREVIATURAS... XIX ÍNDICE DE CONTENIDO... XXIII ÍNDICE DE FIGURAS... XXXI ÍNDICE DE TABLAS... XXXVII INTRODUCCIÓN... 1 1. La mitocondria...
ÍNDICE AGRADECIMIENTOS... VII RESUMEN... XIII RESUM... XV SUMMARY... XVII ABREVIATURAS... XIX ÍNDICE DE CONTENIDO... XXIII ÍNDICE DE FIGURAS... XXXI ÍNDICE DE TABLAS... XXXVII INTRODUCCIÓN... 1 1. La mitocondria...
Novedades de St. Gallen: Diagnóstico y Manejo del cáncer de mama temprano triple negativo
 Novedades de St. Gallen: Diagnóstico y Manejo del cáncer de mama temprano triple negativo Dra. Bettina Müller Oncóloga Médico Instituto Nacional del Cáncer Ref.: Prat A, Perou C: Mammary development meets
Novedades de St. Gallen: Diagnóstico y Manejo del cáncer de mama temprano triple negativo Dra. Bettina Müller Oncóloga Médico Instituto Nacional del Cáncer Ref.: Prat A, Perou C: Mammary development meets
Dr Alejandro Acevedo Gaete Oncólogo Médico Hospital Carlos Van Buren
 Dr Alejandro Acevedo Gaete Oncólogo Médico Hospital Carlos Van Buren Introduccion Perspectiva histórica Principios de Biologia molecular del cáncer colorectal Conceptos de biomarcadores Evidencia clínica
Dr Alejandro Acevedo Gaete Oncólogo Médico Hospital Carlos Van Buren Introduccion Perspectiva histórica Principios de Biologia molecular del cáncer colorectal Conceptos de biomarcadores Evidencia clínica
IDENTIFICACIÓN Y CARACTERIZACIÓN DE LOS MECANISMOS DE TUMORIGÉNESIS MEDIADOS POR FRAGMENTOS CARBOXITERMINALES DE HER2
 IDENTIFICACIÓN Y CARACTERIZACIÓN DE LOS MECANISMOS DE TUMORIGÉNESIS MEDIADOS POR FRAGMENTOS CARBOXITERMINALES DE HER2 Investigador principal: Dr. Joaquín V. Arribas López Hospital Universitari Vall d Hebron
IDENTIFICACIÓN Y CARACTERIZACIÓN DE LOS MECANISMOS DE TUMORIGÉNESIS MEDIADOS POR FRAGMENTOS CARBOXITERMINALES DE HER2 Investigador principal: Dr. Joaquín V. Arribas López Hospital Universitari Vall d Hebron
Factores predictivos. José Palacios. Anatomía Patológica Hospital Universitario Ramón y Cajal Madrid.
 Factores predictivos en cáncer c de mama José Palacios. Anatomía Patológica Hospital Universitario Ramón y Cajal Madrid. CELLULAR TYPES IN MAMMARY GLAND Luminal epithelial CK8, 18, 19 Cadherina-E Myoepithelial
Factores predictivos en cáncer c de mama José Palacios. Anatomía Patológica Hospital Universitario Ramón y Cajal Madrid. CELLULAR TYPES IN MAMMARY GLAND Luminal epithelial CK8, 18, 19 Cadherina-E Myoepithelial
BIOLOGIA TUMORAL CURSO DE ONCOLOGIA MOLECULAR. Dr. José Mordoh. Oncología Molecular, 2011
 BIOLOGIA TUMORAL CURSO DE ONCOLOGIA MOLECULAR Dr. José Mordoh. Oncología Molecular, 2011 Orquesta. Bruno Brudovic EL TUMOR SE COMPORTA COMO UN ORGANISMO AUTONOMO LOCURA, de Francis Hammond Proliferación
BIOLOGIA TUMORAL CURSO DE ONCOLOGIA MOLECULAR Dr. José Mordoh. Oncología Molecular, 2011 Orquesta. Bruno Brudovic EL TUMOR SE COMPORTA COMO UN ORGANISMO AUTONOMO LOCURA, de Francis Hammond Proliferación
Detección Marcadores Moleculares por PCR. Dr. Carlos G. Gonzalez Becuar
 Detección Marcadores Moleculares por PCR Dr. Carlos G. Gonzalez Becuar Virus de Papiloma Humano (VPH) - El CaCu ocupa los primeros lugares por muertes por Cancer en Mexico - Factor principal: VPH tipos
Detección Marcadores Moleculares por PCR Dr. Carlos G. Gonzalez Becuar Virus de Papiloma Humano (VPH) - El CaCu ocupa los primeros lugares por muertes por Cancer en Mexico - Factor principal: VPH tipos
TIPOS HISTOLOGICOS CANCER MAMA
 TIPOS HISTOLOGICOS DE CANCER MAMA WHO HISTOLOGICAL CLASSIFICATION OF TUMORS OF THE BREAST 2003 WHO HISTOLOGICAL CLASSIFICATION OF TUMORS OF THE BREAST 2012 NATURE 10 SUBTIPOS DE CÁNCER DE MAMA "El cáncer
TIPOS HISTOLOGICOS DE CANCER MAMA WHO HISTOLOGICAL CLASSIFICATION OF TUMORS OF THE BREAST 2003 WHO HISTOLOGICAL CLASSIFICATION OF TUMORS OF THE BREAST 2012 NATURE 10 SUBTIPOS DE CÁNCER DE MAMA "El cáncer
La Biología Molecular y la secuenciación masiva abren un nuevo escenario en el abordaje personalizado del cáncer de mama
 Nota de prensa: La Biología Molecular y la secuenciación masiva abren un nuevo escenario en el abordaje personalizado del cáncer de mama CLIPPING DE COBERTURA Enero de 2014 MEDIOS IMPRESOS 17/01/2014 MEDIOS
Nota de prensa: La Biología Molecular y la secuenciación masiva abren un nuevo escenario en el abordaje personalizado del cáncer de mama CLIPPING DE COBERTURA Enero de 2014 MEDIOS IMPRESOS 17/01/2014 MEDIOS
Cáncer de Mama: Inhibidores de vía PI3K/Akt/mTOR, ciclinas, anti-ctla4 y anti PD1. Dr. Claudio Painemal D. Director Médico
 Cáncer de Mama: Inhibidores de vía PI3K/Akt/mTOR, ciclinas, anti-ctla4 y anti PD1 Dr. Claudio Painemal D. Director Médico PI3K/Akt/mTOR Frecuentemente activada en cáncer de mama PIK3CA es el gen más frecuentemente
Cáncer de Mama: Inhibidores de vía PI3K/Akt/mTOR, ciclinas, anti-ctla4 y anti PD1 Dr. Claudio Painemal D. Director Médico PI3K/Akt/mTOR Frecuentemente activada en cáncer de mama PIK3CA es el gen más frecuentemente
Medicina individualizada: Cáncer de mama. Cáncer heredofamiliar. Encarna Adrover CHUA Albacete
 Medicina individualizada: Cáncer de mama. Cáncer heredofamiliar Encarna Adrover CHUA Albacete Bases terapéuticas del cáncer mama Radiología Clínica Comité multidisciplinar Tratamiento de la lesión Tratamiento
Medicina individualizada: Cáncer de mama. Cáncer heredofamiliar Encarna Adrover CHUA Albacete Bases terapéuticas del cáncer mama Radiología Clínica Comité multidisciplinar Tratamiento de la lesión Tratamiento
CANCER DE MAMA DR. MIGUEL SAMEC HOSPITAL DR. TEODORO ALVAREZ (CABA)
") CANCER DE MAMA DR. MIGUEL SAMEC HOSPITAL DR. TEODORO ALVAREZ (CABA) EPIDEMIOLOGIA MAYOR INCIDENCIA ENTRE MUJERES DE 45 A 70 AÑOS PRIMERA CAUSA DE MUERTE EN MUJERES POR CANCER EN ARGENTINA SE PRODUCEN 5400
CANCER DE MAMA DR. MIGUEL SAMEC HOSPITAL DR. TEODORO ALVAREZ (CABA) EPIDEMIOLOGIA MAYOR INCIDENCIA ENTRE MUJERES DE 45 A 70 AÑOS PRIMERA CAUSA DE MUERTE EN MUJERES POR CANCER EN ARGENTINA SE PRODUCEN 5400
GENÉTICA MOLECULAR. Unidad 1: Introducción a la genética molecular
 GENÉTICA MOLECULAR CONTENIDOS CONCEPTUALES COMPETENCIAS Unidad 1: Introducción a la genética molecular Concepto de genética molecular y aplicaciones a diferentes ramas de la ciencia. Dogma central de la
GENÉTICA MOLECULAR CONTENIDOS CONCEPTUALES COMPETENCIAS Unidad 1: Introducción a la genética molecular Concepto de genética molecular y aplicaciones a diferentes ramas de la ciencia. Dogma central de la
PREVENCION DEL CÁNCER GINECOLÓGICO CÁNCER DE MAMA DRA. RAMÍREZ MEDINA SERVICIO DE GINECOLOGÍA Y OBSTETRICIA UNIDAD DE MAMA
 PREVENCION DEL CÁNCER GINECOLÓGICO CÁNCER DE MAMA DRA. RAMÍREZ MEDINA SERVICIO DE GINECOLOGÍA Y OBSTETRICIA UNIDAD DE MAMA QUÉ ES EL CÁNCER DE MAMA? Es el tumor maligno que se origina en las células de
PREVENCION DEL CÁNCER GINECOLÓGICO CÁNCER DE MAMA DRA. RAMÍREZ MEDINA SERVICIO DE GINECOLOGÍA Y OBSTETRICIA UNIDAD DE MAMA QUÉ ES EL CÁNCER DE MAMA? Es el tumor maligno que se origina en las células de
IMPACTO DIAGNÓSTICO DE LOS AVANCES MOLECULARES EN CÁNCER DE MAMA. Papel de la transición epitelio mesénquima en el cáncer de mama
 IMPACTO DIAGNÓSTICO DE LOS AVANCES MOLECULARES EN CÁNCER DE MAMA. Papel de la transición epitelio mesénquima en el cáncer de mama Enrique Lerma. Servicio de Anatomía Patológica Hospital de la Santa Creu
IMPACTO DIAGNÓSTICO DE LOS AVANCES MOLECULARES EN CÁNCER DE MAMA. Papel de la transición epitelio mesénquima en el cáncer de mama Enrique Lerma. Servicio de Anatomía Patológica Hospital de la Santa Creu
Blga. María Leticia Amésquita Cárdenas. GENETICA MEDICA Departamento Morfología Humana. Facultad de Medicina Universidad Nacional de Trujillo
 Blga. María Leticia Amésquita Cárdenas GENETICA MEDICA Departamento Morfología Humana. Facultad de Medicina Universidad Nacional de Trujillo Cáncer Enfermedad neoplásica con transformación de las células
Blga. María Leticia Amésquita Cárdenas GENETICA MEDICA Departamento Morfología Humana. Facultad de Medicina Universidad Nacional de Trujillo Cáncer Enfermedad neoplásica con transformación de las células
Cáncer de Mama Resistencia a tratamientos anti- HER2 en pacientes con Cáncer de mama.
 Cáncer de Mama Resistencia a tratamientos anti- HER2 en pacientes con Cáncer de mama. Dr. Atanasio Pandiella, Centro Investigación Cáncer, Salamanca Dr. Alberto Ocaña, Complejo Hosp. Univeristario de Albacete
Cáncer de Mama Resistencia a tratamientos anti- HER2 en pacientes con Cáncer de mama. Dr. Atanasio Pandiella, Centro Investigación Cáncer, Salamanca Dr. Alberto Ocaña, Complejo Hosp. Univeristario de Albacete
Descripción CLASES MAGISTRALES, TALLERES PRÁCTICOS DE CITOLOGÍA Y PRESENTACIÓN DE CASOS CLÍNICOS
 CURSO DE POSGRADO EN ONCOLOGÍA VETERINARIA De lo molecular a lo clínico Facultad de Ciencias Agropecuarias Universidad Católica de Córdoba Cupo MÍNIMO 10, MÁXIMO 30 ALUMNOS Descripción CLASES MAGISTRALES,
CURSO DE POSGRADO EN ONCOLOGÍA VETERINARIA De lo molecular a lo clínico Facultad de Ciencias Agropecuarias Universidad Católica de Córdoba Cupo MÍNIMO 10, MÁXIMO 30 ALUMNOS Descripción CLASES MAGISTRALES,
28-01-2012 BASES GENÉTICAS DEL CÁNCER. Ciclo Celular. Diferenciación
 Ciclo Celular BASES GENÉTICAS DEL CÁNCER Las células proliferan se diferencian y mueren desarrollo y mantenimiento de tejidos y órganos Proceso controlado por complejo mecanismo señales intracelulares
Ciclo Celular BASES GENÉTICAS DEL CÁNCER Las células proliferan se diferencian y mueren desarrollo y mantenimiento de tejidos y órganos Proceso controlado por complejo mecanismo señales intracelulares
IDENTIFICACIÓN DE NUEVAS DIANAS TERAPÉUTICAS EN TUMORES GERMINALES REFRACTARIOS
 IDENTIFICACIÓN DE NUEVAS DIANAS TERAPÉUTICAS EN TUMORES GERMINALES REFRACTARIOS Investigadores principales: Dr. Francesc Viñals Canals Facultat de Medicina CSUB Dr. Fco. Javier García del Muro Solans Institut
IDENTIFICACIÓN DE NUEVAS DIANAS TERAPÉUTICAS EN TUMORES GERMINALES REFRACTARIOS Investigadores principales: Dr. Francesc Viñals Canals Facultat de Medicina CSUB Dr. Fco. Javier García del Muro Solans Institut
Selección de pacientes
 2 Selección de pacientes MENSAJES CLAVE Aproximadamente entre el 20% y el 30% de las pacientes con cáncer de mama presentan tumores HER2 positivos. Hasta un 24% de los tumores con receptores de estrógenos
2 Selección de pacientes MENSAJES CLAVE Aproximadamente entre el 20% y el 30% de las pacientes con cáncer de mama presentan tumores HER2 positivos. Hasta un 24% de los tumores con receptores de estrógenos
PharmaMar presentara los últimos datos obtenidos con sus compuestos PM1183 y plitidepsina en el Congreso Anual AACR 2015
 PharmaMar presentara los últimos datos obtenidos con sus compuestos PM1183 y plitidepsina en el Congreso Anual AACR 2015 El compuesto PM1183 en combinación con doxorrubicina produce un efecto antitumoral
PharmaMar presentara los últimos datos obtenidos con sus compuestos PM1183 y plitidepsina en el Congreso Anual AACR 2015 El compuesto PM1183 en combinación con doxorrubicina produce un efecto antitumoral
OBJETIVOS DEL TRATAMIENTO QUIRÚRGICO. -Resección del tejido tumoral con márgenes oncológicos adecuados. - Mejor resultado cosmético posible
 MANEJO QUIRÚRGICO DEL CÁNCER DE MAMA RECUERDO HISTÓRICO Siglo XV a.c. Papiro de Ebersdescribe el tratamiento de los tumores de mama con hierro o con fuego Siglo I a.c. Siglo I d.c. Siglo XVI Siglo XVIII
MANEJO QUIRÚRGICO DEL CÁNCER DE MAMA RECUERDO HISTÓRICO Siglo XV a.c. Papiro de Ebersdescribe el tratamiento de los tumores de mama con hierro o con fuego Siglo I a.c. Siglo I d.c. Siglo XVI Siglo XVIII
ESTUDIOS GENÉTICOS EN CÁNCER HEREDITARIO
 ESTUDIOS GENÉTICOS EN CÁNCER HEREDITARIO ENTIDAD GEN SÍNDROME ASOCIADO BRCA1 y BRCA2 Ca de mama/ovario Ca de mama/ovario TP53 Li Fraumeni PTEN Enfermedad de Cowden STK11 Sindrome Peutz-Jeghers Ca de mama
ESTUDIOS GENÉTICOS EN CÁNCER HEREDITARIO ENTIDAD GEN SÍNDROME ASOCIADO BRCA1 y BRCA2 Ca de mama/ovario Ca de mama/ovario TP53 Li Fraumeni PTEN Enfermedad de Cowden STK11 Sindrome Peutz-Jeghers Ca de mama
Enfermedades genéticas
 Enfermedades genéticas Desórdenes genéticos Desórdenes genéticos Mutaciones Rearreglos genómicos Variación en número de cromosomas Congénitos Línea germinal somáticos Aparecer en algún momento post nacimiento
Enfermedades genéticas Desórdenes genéticos Desórdenes genéticos Mutaciones Rearreglos genómicos Variación en número de cromosomas Congénitos Línea germinal somáticos Aparecer en algún momento post nacimiento
CLUB DE PATOLOGÍA DE LA MAMA
 CLUB DE PATOLOGÍA DE LA MAMA Aprendiendo de nuestros errores Laura López Vilaró CASO CLÍNICO Mujer 66 años sin antecedentes patológicos destacables Nódulo retro-areolar de mama derecha en mamografía del
CLUB DE PATOLOGÍA DE LA MAMA Aprendiendo de nuestros errores Laura López Vilaró CASO CLÍNICO Mujer 66 años sin antecedentes patológicos destacables Nódulo retro-areolar de mama derecha en mamografía del
Proyecto CENIT. III Conferencia Anual de las Plataformas Tecnológicas de Investigación Biomédica
 Proyecto CENIT III Conferencia Anual de las Plataformas Tecnológicas de Investigación Biomédica Miguel Aracil (Coordinador científico del Consorcio ONCOLOGICA) Barcelona, 23-24 Febrero 2010 Consorcio ONCOLOGICA
Proyecto CENIT III Conferencia Anual de las Plataformas Tecnológicas de Investigación Biomédica Miguel Aracil (Coordinador científico del Consorcio ONCOLOGICA) Barcelona, 23-24 Febrero 2010 Consorcio ONCOLOGICA
EL EQUIPO MULTIDISCIPLINAR XXVI CONGRESO DE LA SEAP/IAP PAPEL DEL PATOLOGO EN LOS TUMORES DEL APARATO DIGESTIVO BIOMARCADORES PREDICTIVOS
 XXVI CONGRESO DE LA SEAP/IAP PAPEL DEL PATOLOGO EN LOS TUMORES DEL APARATO DIGESTIVO ACTUALIZACION EN BIOMARCADORES Y GUIA DE RECOMENDACIONES EN TUMORES DEL APARATO DIGESTIVO # DIAGNOSTICO # APORTACION
XXVI CONGRESO DE LA SEAP/IAP PAPEL DEL PATOLOGO EN LOS TUMORES DEL APARATO DIGESTIVO ACTUALIZACION EN BIOMARCADORES Y GUIA DE RECOMENDACIONES EN TUMORES DEL APARATO DIGESTIVO # DIAGNOSTICO # APORTACION
CÓMO SE PUEDE DETERMINAR LA SECUENCIA DEL DNA A PARTIR DE UNA PROTEÍNA? DR. MANUEL E. AQUINO
 Alianza para el Aprendizaje de Ciencias y Matemáticas CÓMO SE PUEDE DETERMINAR LA SECUENCIA DEL DNA A PARTIR DE UNA PROTEÍNA? DR. MANUEL E. AQUINO GUÍA DEL MAESTRO ESTÁNDARES ATENDIDOS: 1. LA NATURALEZA
Alianza para el Aprendizaje de Ciencias y Matemáticas CÓMO SE PUEDE DETERMINAR LA SECUENCIA DEL DNA A PARTIR DE UNA PROTEÍNA? DR. MANUEL E. AQUINO GUÍA DEL MAESTRO ESTÁNDARES ATENDIDOS: 1. LA NATURALEZA
Dra. Mónica Adriana Carrera Álvarez
 Dra. Mónica Adriana Carrera Álvarez INCIDENCIA Y EPIDEMIOLOGIA: Actualmente ha habido una disminución del 30 al 20% en la tasa de mortalidad. Mujeres blancas más frecuente Actualmente tendencia ha aumentado
Dra. Mónica Adriana Carrera Álvarez INCIDENCIA Y EPIDEMIOLOGIA: Actualmente ha habido una disminución del 30 al 20% en la tasa de mortalidad. Mujeres blancas más frecuente Actualmente tendencia ha aumentado
OPTIMIZACIÓN TRATAMIENTO ANTI-EGFR. Ruth Vera Oncología Médica
 OPTIMIZACIÓN TRATAMIENTO ANTI-EGFR Ruth Vera Oncología Médica OPTIMIZACIÓN TRATAMIENTO anti-egfr OPTIMIZAR quiere decir: Buscar los mejores resultados Planificar una actividad para obtener los mejores
OPTIMIZACIÓN TRATAMIENTO ANTI-EGFR Ruth Vera Oncología Médica OPTIMIZACIÓN TRATAMIENTO anti-egfr OPTIMIZAR quiere decir: Buscar los mejores resultados Planificar una actividad para obtener los mejores
PROGRAMA DE DOCTORADO EN CIENCIAS BIOLÓGICAS MENCIÓN CIENCIAS FISIOLÓGICAS. : Dr. Ricardo Moreno y Dr. Nelson Barrera. : Lunes - Miércoles - Viernes.
 PROGRAMA DE DOCTORADO EN CIENCIAS BIOLÓGICAS MENCIÓN CIENCIAS FISIOLÓGICAS I. ANTECEDENTES GENERALES Nombre del Curso Sigla Carácter : COMUNICACIÓN CELULAR. : BIO4141 : Mínimo. Créditos : 15. Profesor
PROGRAMA DE DOCTORADO EN CIENCIAS BIOLÓGICAS MENCIÓN CIENCIAS FISIOLÓGICAS I. ANTECEDENTES GENERALES Nombre del Curso Sigla Carácter : COMUNICACIÓN CELULAR. : BIO4141 : Mínimo. Créditos : 15. Profesor
Acaban de diagnosticarle cáncer de
 Acaban de diagnosticarle cáncer de mama en estadío temprano? Laurie L. Escritora y antropóloga. Diagnosticada con cáncer de mama invasivo en el 2005. Una guía educativa elaborada por Genomic Health Esta
Acaban de diagnosticarle cáncer de mama en estadío temprano? Laurie L. Escritora y antropóloga. Diagnosticada con cáncer de mama invasivo en el 2005. Una guía educativa elaborada por Genomic Health Esta
Cáncer de Mama Opciones Terapéuticas SIGLO XXI
 Cáncer de Mama Opciones Terapéuticas SIGLO XXI Prof Jaime Sanz Ortiz Servicio 29 Marzo de 2010 Oncología Médica y Cuidados Paliativos. HUMV Jaime Sanz Ortiz 1 LOS MUNDOS DEL CÁNCER 1970-1990 Oncología
Cáncer de Mama Opciones Terapéuticas SIGLO XXI Prof Jaime Sanz Ortiz Servicio 29 Marzo de 2010 Oncología Médica y Cuidados Paliativos. HUMV Jaime Sanz Ortiz 1 LOS MUNDOS DEL CÁNCER 1970-1990 Oncología
Investigadora principal: Dra. Rosanna Paciucci Barzanti Hospital Universitari Vall d Hebron Duración: 3 años
 CARACTERIZACIÓN FUNCIONAL DEL COMPLEJO PTOV1-FLOTILINA-1 EN LA MODULACIÓN DE LA PROLIFERACIÓN DE CÉLULAS CANCEROSAS: CONTROL MITÓTICO E INTERACCIÓN CON LA VÍA NOTCH Investigadora principal: Dra. Rosanna
CARACTERIZACIÓN FUNCIONAL DEL COMPLEJO PTOV1-FLOTILINA-1 EN LA MODULACIÓN DE LA PROLIFERACIÓN DE CÉLULAS CANCEROSAS: CONTROL MITÓTICO E INTERACCIÓN CON LA VÍA NOTCH Investigadora principal: Dra. Rosanna
BIOQUIMICA Y GENETICA MOLECULAR APLICADA A LA VETERINARIA
 BIOQUIMICA Y GENETICA MOLECULAR APLICADA A LA VETERINARIA PROFESORADO CURSO 1994-1995: Dr. J. Ariño (Unidad de Bioquímica) Dra. F. Bosch (Unidad de Bioquímica) Dr. A. Sánchez (Unidad de Genética y Mejora)
BIOQUIMICA Y GENETICA MOLECULAR APLICADA A LA VETERINARIA PROFESORADO CURSO 1994-1995: Dr. J. Ariño (Unidad de Bioquímica) Dra. F. Bosch (Unidad de Bioquímica) Dr. A. Sánchez (Unidad de Genética y Mejora)
Biología (54208) Profesora: Dra. Marlys Campos
 Profesora: Dra. Marlys Campos") Biología (54208) Profesora: Dra. Marlys Campos 2-2016 Profesora Teoría: Dra. Marlys Campos C. http://www.udesantiagovirtual.cl/moodle2/ Teoría: Lunes (13:50-15:20) Jueves (9:40-11:10) Laboratorio: Lunes
Biología (54208) Profesora: Dra. Marlys Campos 2-2016 Profesora Teoría: Dra. Marlys Campos C. http://www.udesantiagovirtual.cl/moodle2/ Teoría: Lunes (13:50-15:20) Jueves (9:40-11:10) Laboratorio: Lunes
Precisión en la identificación de los tumores Her2
 Precisión en la identificación de los tumores Her2 CLASIFICACION MOLECULAR DEL CANCER DE MAMA SUBTIPOS DE CARCINOMAS MAMARIOS DETERMINADOS POR PERFILES DE EXPRESION GENETICA APROXIMACIÓN INMUNOFENOTÍPICA
Precisión en la identificación de los tumores Her2 CLASIFICACION MOLECULAR DEL CANCER DE MAMA SUBTIPOS DE CARCINOMAS MAMARIOS DETERMINADOS POR PERFILES DE EXPRESION GENETICA APROXIMACIÓN INMUNOFENOTÍPICA
Estudio de la función de la variante de histona macroh2a2 en el desarrollo de la retina del pez cebra
 UNIVERSIDAD ANDRÉS BELLO FACULTAD DE CIENCIAS BIOLOGICAS DOCTORADO DE BIOCIENCIAS MOLECULARES Estudio de la función de la variante de histona macroh2a2 en el desarrollo de la retina del pez cebra Tesis
UNIVERSIDAD ANDRÉS BELLO FACULTAD DE CIENCIAS BIOLOGICAS DOCTORADO DE BIOCIENCIAS MOLECULARES Estudio de la función de la variante de histona macroh2a2 en el desarrollo de la retina del pez cebra Tesis
Discapacidad Intelectual 360º
 Discapacidad Intelectual 360º Te garantizamos la máxima utilidad clínica El mejor servicio del mercado para diagnosticar Discapacidad Intelectual La frecuencia de este trastorno en menores de 18 años se
Discapacidad Intelectual 360º Te garantizamos la máxima utilidad clínica El mejor servicio del mercado para diagnosticar Discapacidad Intelectual La frecuencia de este trastorno en menores de 18 años se
Oncología Molecular MBMM 2013. Bioq. María Paula Roberti Bioq. Juan Martín Arriaga
 Oncología Molecular MBMM 2013 Bioq. María Paula Roberti Bioq. Juan Martín Arriaga El cáncer de mama La Agencia Internacional de Investigación sobre el Cáncer ha estimado que durante el año 2008 en Argentina
Oncología Molecular MBMM 2013 Bioq. María Paula Roberti Bioq. Juan Martín Arriaga El cáncer de mama La Agencia Internacional de Investigación sobre el Cáncer ha estimado que durante el año 2008 en Argentina
7.2 Análisis de Metilación en Muestras de Tejido
 VII. DISCUSIÓN 7.1 Evaluación del Método de Análisis de Metilación Específica y Cuantitativa de Múltiples Genes por PCR en Tiempo Real (QM-MSP) en Líneas Celulares y Muestras de Tejido Microdisectadas
VII. DISCUSIÓN 7.1 Evaluación del Método de Análisis de Metilación Específica y Cuantitativa de Múltiples Genes por PCR en Tiempo Real (QM-MSP) en Líneas Celulares y Muestras de Tejido Microdisectadas
Dr. Horacio Decanini Arcaute Anatomopatólogo
 Dr. Horacio Decanini Arcaute Anatomopatólogo Consenso general Introducción: La sobre- expresión del gen Her / 2 neu se encuentra Asociada con comportamiento agresivo y es considerada Como factor predictivo
Dr. Horacio Decanini Arcaute Anatomopatólogo Consenso general Introducción: La sobre- expresión del gen Her / 2 neu se encuentra Asociada con comportamiento agresivo y es considerada Como factor predictivo
Qué es la Genética? La Genética es el estudio de la Herencia Biológica GENETICA GENETICA HUMANA caracteres biológicos GENETICA MEDICA
 Qué es la Genética? La Genética es el estudio de la Herencia Biológica Estudio de la unidad de la herencia: GEN Los mecanismos y patrones de herencia de los caracteres biológicos La variabilidad entre
Qué es la Genética? La Genética es el estudio de la Herencia Biológica Estudio de la unidad de la herencia: GEN Los mecanismos y patrones de herencia de los caracteres biológicos La variabilidad entre
Patologías asociadas a defectos en la reparación del DNA. Xeroderma pigmentosa (XP) Ataxia-telangiectasia (A-T)
 Ataxia-telangiectasia (A-T)") Patologías asociadas a defectos en la reparación del DNA Xeroderma pigmentosa (XP) Ataxia-telangiectasia (A-T) Defectos en los mecanismos de reparación del DNA se traducen en una gran probabilidad de desarrollar
Patologías asociadas a defectos en la reparación del DNA Xeroderma pigmentosa (XP) Ataxia-telangiectasia (A-T) Defectos en los mecanismos de reparación del DNA se traducen en una gran probabilidad de desarrollar
Hallazgos anatomopatológicos en pacientes con una punción aspiración con aguja fina o biopsia con aguja gruesa en el tórax.
 Hallazgos anatomopatológicos en pacientes con una punción aspiración con aguja fina o biopsia con aguja gruesa en el tórax. Poster no.: S-0623 Congreso: SERAM 2014 Tipo del póster: Presentación Electrónica
Hallazgos anatomopatológicos en pacientes con una punción aspiración con aguja fina o biopsia con aguja gruesa en el tórax. Poster no.: S-0623 Congreso: SERAM 2014 Tipo del póster: Presentación Electrónica
CARCINOMA DUCTAL IN SITU E INVASOR
 CARCINOMA DUCTAL IN SITU E INVASOR DRA. ALEJANDRA ZÁRATE OSORNO HOSPITAL ESPAÑOL DE MEXICO CARCINOMA DUCTAL IN SITU PROLIFERACIÓN MALIGNA DE CÉLULAS EPITELIALES DENTRO DE LOS DUCTOS, SIN EVIDENCIA DE INVASIÓN
CARCINOMA DUCTAL IN SITU E INVASOR DRA. ALEJANDRA ZÁRATE OSORNO HOSPITAL ESPAÑOL DE MEXICO CARCINOMA DUCTAL IN SITU PROLIFERACIÓN MALIGNA DE CÉLULAS EPITELIALES DENTRO DE LOS DUCTOS, SIN EVIDENCIA DE INVASIÓN
EPIDEMIOLOGÍA. Sexo Femenino (entre los 10 primeros cánceres) Frecuencia de CT según distribución por sexo
 Frecuencia de CT según distribución por sexo") EPIDEMIOLOGÍA El cáncer de tiroides es el tumor endocrino mas frecuente (90%) y representa el 1% de todas las neoplasias malignas del ser humano. Aproximadamente un 10% de la población desarrollará un
EPIDEMIOLOGÍA El cáncer de tiroides es el tumor endocrino mas frecuente (90%) y representa el 1% de todas las neoplasias malignas del ser humano. Aproximadamente un 10% de la población desarrollará un
DEDICATORIAS. A Dios, Esta tesis se la dedicamos a Dios quién supo guiarnos por el buen camino,
 DEDICATORIAS A Dios, Esta tesis se la dedicamos a Dios quién supo guiarnos por el buen camino, dándonos las fuerzas para seguir adelante y no desmayar en los problemas que se presentaban, enseñándonos
DEDICATORIAS A Dios, Esta tesis se la dedicamos a Dios quién supo guiarnos por el buen camino, dándonos las fuerzas para seguir adelante y no desmayar en los problemas que se presentaban, enseñándonos
ABREVIATURAS... XI I. INTRODUCCIÓN EL TOMATE... 3
 ÍNDICE ABREVIATURAS... XI I. INTRODUCCIÓN... 1 1 EL TOMATE... 3 1.1 Taxonomía... 3 1.2 Características generales... 3 1.3 La flor... 4 1.4 Características del fruto... 5 2 CUAJADO Y DESARROLLO DEL FRUTO...
ÍNDICE ABREVIATURAS... XI I. INTRODUCCIÓN... 1 1 EL TOMATE... 3 1.1 Taxonomía... 3 1.2 Características generales... 3 1.3 La flor... 4 1.4 Características del fruto... 5 2 CUAJADO Y DESARROLLO DEL FRUTO...
Análisis de la coestimulación vía CD28 en células linfoides de pacientes infectados. con el virus de la Hepatitis C RESUMEN
 Análisis de la coestimulación vía CD28 en células linfoides de pacientes infectados con el virus de la Hepatitis C RESUMEN La infección por el virus de la hepatitis C (VHC) afecta a más de 170 millones
Análisis de la coestimulación vía CD28 en células linfoides de pacientes infectados con el virus de la Hepatitis C RESUMEN La infección por el virus de la hepatitis C (VHC) afecta a más de 170 millones
Perfiles de expresión génica y diagnóstico/pronóstico en cáncer. Introducción 6/1/2010. Meritxell Pelicano Esqueta. Genómica y proteómica
 Perfiles de expresión génica y diagnóstico/pronóstico en cáncer Meritxell Pelicano Esqueta Genómica y proteómica Curso 2009/2010 Introducción Expresión génica: Proceso por el cual los organismos transforman
Perfiles de expresión génica y diagnóstico/pronóstico en cáncer Meritxell Pelicano Esqueta Genómica y proteómica Curso 2009/2010 Introducción Expresión génica: Proceso por el cual los organismos transforman
BIOPSIAS DE PRÓSTATA POR PUNCIÓN (II)
") IV CONGRESO VIRTUAL HISPANO AMERICANO DE ANATOMÍA PATOLÓGICA CONTENIDO Caso anterior Caso siguiente Título preliminar Presentación Imágenes Resumen Material Resultados Diagnósticos PDF BIOPSIAS DE PRÓSTATA
IV CONGRESO VIRTUAL HISPANO AMERICANO DE ANATOMÍA PATOLÓGICA CONTENIDO Caso anterior Caso siguiente Título preliminar Presentación Imágenes Resumen Material Resultados Diagnósticos PDF BIOPSIAS DE PRÓSTATA
Tumor phyllodes. Erika Fonseca Chimá Residente de obstetricia Universidad de Antioquia. Asesor: Dr. German Garcia
 Tumor phyllodes Erika Fonseca Chimá Residente de obstetricia Universidad de Antioquia Asesor: Dr. German Garcia Carcinoma: mayoría de tumores malignos de mama Sarcomas primarios de mama: tejido mesenquimal
Tumor phyllodes Erika Fonseca Chimá Residente de obstetricia Universidad de Antioquia Asesor: Dr. German Garcia Carcinoma: mayoría de tumores malignos de mama Sarcomas primarios de mama: tejido mesenquimal
ASOCIACION DEL P53 A LOS PRINCIPALES FACTORES PRONOSTICOS EN PACIENTES CON MELANOMA CUTANEO MALIGNO.
 ASOCIACION DEL P53 A LOS PRINCIPALES FACTORES PRONOSTICOS EN PACIENTES CON MELANOMA CUTANEO MALIGNO. Dra. MIRLA MIRANDA. Dr. ELIECER PAYARES. Dr. ALDO REIGOSA. INTRODUCCION. - El Melanoma cutáneo: alta
ASOCIACION DEL P53 A LOS PRINCIPALES FACTORES PRONOSTICOS EN PACIENTES CON MELANOMA CUTANEO MALIGNO. Dra. MIRLA MIRANDA. Dr. ELIECER PAYARES. Dr. ALDO REIGOSA. INTRODUCCION. - El Melanoma cutáneo: alta
CLUB DE PATOLOGÍA MAMARIA
 CLUB DE PATOLOGÍA MAMARIA Congreso Nacional de la SEAP-IAP Cádiz, mayo 2013 Rosario Granados Caso Clínico Mujer de 60 años con nódulo de 1,5 cm en CSE de mama derecha. Tumorectomía. Diagnóstico
CLUB DE PATOLOGÍA MAMARIA Congreso Nacional de la SEAP-IAP Cádiz, mayo 2013 Rosario Granados Caso Clínico Mujer de 60 años con nódulo de 1,5 cm en CSE de mama derecha. Tumorectomía. Diagnóstico
PLANIFICACIÓN DE ASIGNATURA
 PLANIFICACIÓN DE ASIGNATURA I IDENTIFICACION GENERAL DE LA ASIGNATURA CARRERA BIOQUIMICA DEPARTAMENTO BIOLOGIA ASIGNATURA BIOLOGIA MOLECULAR II CÓDIGO 1681 PRERREQUISITOS Biología Molecular I CREDITOS
PLANIFICACIÓN DE ASIGNATURA I IDENTIFICACION GENERAL DE LA ASIGNATURA CARRERA BIOQUIMICA DEPARTAMENTO BIOLOGIA ASIGNATURA BIOLOGIA MOLECULAR II CÓDIGO 1681 PRERREQUISITOS Biología Molecular I CREDITOS
Problemas Diagnosticos en Lesiones Foliculares de la Glandula Tiroides. Saul Suster, M.D. Medical College of Wisconsin Milwaukee, WI, USA
 Problemas Diagnosticos en Lesiones Foliculares de la Glandula Tiroides Saul Suster, M.D. Medical College of Wisconsin Milwaukee, WI, USA Lesiones Foliculares Benignas de Tiroides que Pueden Confundirse
Problemas Diagnosticos en Lesiones Foliculares de la Glandula Tiroides Saul Suster, M.D. Medical College of Wisconsin Milwaukee, WI, USA Lesiones Foliculares Benignas de Tiroides que Pueden Confundirse
Anticuerpos Monoclonales contra el Cáncer de mama
 Anticuerpos Monoclonales contra el Cáncer de mama Erick Ovando a la Biotecnología Departamento de Química Universidad Técnica Federico Santa María Valparaíso, 05 de Diciembre de 2006 1 Estadísticas de
Anticuerpos Monoclonales contra el Cáncer de mama Erick Ovando a la Biotecnología Departamento de Química Universidad Técnica Federico Santa María Valparaíso, 05 de Diciembre de 2006 1 Estadísticas de
Acaban de diagnosticarle cáncer de
 Acaban de diagnosticarle cáncer de mama en fase inicial? Laurie L. Escritora y antropóloga. Diagnosticada con cáncer de mama invasivo en el 2005. Una guía educativa elaborada por Genomic Health Esta guía
Acaban de diagnosticarle cáncer de mama en fase inicial? Laurie L. Escritora y antropóloga. Diagnosticada con cáncer de mama invasivo en el 2005. Una guía educativa elaborada por Genomic Health Esta guía
Bases moleculares y estudios genéticos
 II Jornada Genética Clínica para Médicos de Familia Bases moleculares y estudios genéticos María García-Barcina Unidad de Genética del H. Universitario de Basurto Osakidetza Sede SVMFIC Valencia, 2 de
II Jornada Genética Clínica para Médicos de Familia Bases moleculares y estudios genéticos María García-Barcina Unidad de Genética del H. Universitario de Basurto Osakidetza Sede SVMFIC Valencia, 2 de
Efectos biológicos de las quinasas VRK en cáncer de mama en función de los fenotipos HER2/erbB2 y Receptor de estrógeno
 Convocatoria de Ayudas a la Investigación en Cáncer de Mama Fundación Sandra Ibarra de Solidaridad frente al Cáncer MEMORIA FINAL DEL PROYECTO Efectos biológicos de las quinasas VRK en cáncer de mama en
Convocatoria de Ayudas a la Investigación en Cáncer de Mama Fundación Sandra Ibarra de Solidaridad frente al Cáncer MEMORIA FINAL DEL PROYECTO Efectos biológicos de las quinasas VRK en cáncer de mama en
ENFERMEDADES HEREDITARIAS EN ANIMALES que son las enfermedades hereditarias?
 ENFERMEDADES HEREDITARIAS EN ANIMALES que son las enfermedades hereditarias? Las enfermedades hereditarias son aquellas que se transmiten mediante el proceso de la herencia, es decir de los progenitores
ENFERMEDADES HEREDITARIAS EN ANIMALES que son las enfermedades hereditarias? Las enfermedades hereditarias son aquellas que se transmiten mediante el proceso de la herencia, es decir de los progenitores
EN MUJERES CÁNCER DE MAMA
 JUNIO 2016 SCOR inform CÁNCER DE MAMA EN MUJERES DELPHINE LABOJKA Responsable de Métodos y Procesos Dr PATRICK MALAMUD Oncólogo, consejero médico Dr XUÂN-VIÊT PHAM Reumatólogo, consejero médico Antecedentes
JUNIO 2016 SCOR inform CÁNCER DE MAMA EN MUJERES DELPHINE LABOJKA Responsable de Métodos y Procesos Dr PATRICK MALAMUD Oncólogo, consejero médico Dr XUÂN-VIÊT PHAM Reumatólogo, consejero médico Antecedentes
CITOLOGÍA A POR PUNCIÓN MAMARIA CÉSAR LACRUZ PELEA
 CITOLOGÍA A POR PUNCIÓN MAMARIA CÉSAR LACRUZ PELEA PUNCIÓN N MAMARIA Martin y Ellis (1930s). Memorial Hospital. NY. Dr. Hayes Martin Amplia experiencia mundial en el diagnóstico de nódulos n mediante punción
CITOLOGÍA A POR PUNCIÓN MAMARIA CÉSAR LACRUZ PELEA PUNCIÓN N MAMARIA Martin y Ellis (1930s). Memorial Hospital. NY. Dr. Hayes Martin Amplia experiencia mundial en el diagnóstico de nódulos n mediante punción
RECOMENDACIONES PARA HER 2 IHQ EN CANCER DE MAMA. Dra. Valeria Cornejo C. Hospital Clínico San Borja Arriarán
 RECOMENDACIONES PARA HER 2 IHQ EN CANCER DE MAMA Dra. Valeria Cornejo C. Hospital Clínico San Borja Arriarán Introducción Realización de grandes estudios que evalúan efectividad de las nuevas drogas target
RECOMENDACIONES PARA HER 2 IHQ EN CANCER DE MAMA Dra. Valeria Cornejo C. Hospital Clínico San Borja Arriarán Introducción Realización de grandes estudios que evalúan efectividad de las nuevas drogas target
Son iguales todos los carcinomas de mama HER2+?
 Son iguales todos los carcinomas de mama HER2+? Federico Rojo IIS-Fundación Jiménez Díaz IMIM-Hospital del Mar Curso Corto de Patología Mamaria Nuevos Fenotipos del Cáncer de Mama Nuevos Problemas para
Son iguales todos los carcinomas de mama HER2+? Federico Rojo IIS-Fundación Jiménez Díaz IMIM-Hospital del Mar Curso Corto de Patología Mamaria Nuevos Fenotipos del Cáncer de Mama Nuevos Problemas para
IMPACTO CLINICO DE LA INMUNOFENOTIPIFICION CELULAR IV: Ciclo celular en estados neoplásicos y preneoplásicos. T.M. MSC JUAN LUIS CASTILLO N.
 IMPACTO CLINICO DE LA INMUNOFENOTIPIFICION CELULAR IV: Ciclo celular en estados neoplásicos y preneoplásicos. T.M. MSC JUAN LUIS CASTILLO N. Centro de Diagnóstico Onco-inmunológicoLtda. Laboratorio de
IMPACTO CLINICO DE LA INMUNOFENOTIPIFICION CELULAR IV: Ciclo celular en estados neoplásicos y preneoplásicos. T.M. MSC JUAN LUIS CASTILLO N. Centro de Diagnóstico Onco-inmunológicoLtda. Laboratorio de
LA CITOLOGIA EN EL DIAGNÓSTICO DE LOS TUMORES NEUROENDOCRINOS
 LA CITOLOGIA EN EL DIAGNÓSTICO DE LOS TUMORES NEUROENDOCRINOS Diego Martinez Parra UGC Intercentros de Anatomia Patológica Bahia de Cádiz (Hospital Universitario Puerta del Mar) TUMORES NEUROENDOCRINOS
LA CITOLOGIA EN EL DIAGNÓSTICO DE LOS TUMORES NEUROENDOCRINOS Diego Martinez Parra UGC Intercentros de Anatomia Patológica Bahia de Cádiz (Hospital Universitario Puerta del Mar) TUMORES NEUROENDOCRINOS
Clínica de Enfermedades Infecciosas Hospital Roosevelt Febrero 2016
 Clínica de Enfermedades Infecciosas Hospital Roosevelt Febrero 2016 Requiere de evaluación: Clínica: Signos y Síntomas (la mayoría asintomáticos) Bioquímica Serológica Histológica Virológica Determinar
Clínica de Enfermedades Infecciosas Hospital Roosevelt Febrero 2016 Requiere de evaluación: Clínica: Signos y Síntomas (la mayoría asintomáticos) Bioquímica Serológica Histológica Virológica Determinar
PROGRAMA ANALÍTICO UNIDAD Nº 1: La Universidad. Objetivos Específicos: Contenidos: Universidad: concepto. UNNE: Facultad de Odontología.
 PROGRAMA ANALÍTICO UNIDAD Nº 1: La Universidad. Conocer el ámbito universitario y de la unidad académica Identificar las funciones y las estructuras de gobierno de la UNNE y de Facultad de Odontología
PROGRAMA ANALÍTICO UNIDAD Nº 1: La Universidad. Conocer el ámbito universitario y de la unidad académica Identificar las funciones y las estructuras de gobierno de la UNNE y de Facultad de Odontología
Dra. Dulce María Arreguín Porras Servicio de Infectología Hospital General de México Dr. Eduardo Liceaga
 Dra. Dulce María Arreguín Porras Servicio de Infectología Hospital General de México Dr. Eduardo Liceaga Neoplasias definitorias de SIDA Neoplasias no definitorias de SIDA El grado de inmunosupresión manifestado
Dra. Dulce María Arreguín Porras Servicio de Infectología Hospital General de México Dr. Eduardo Liceaga Neoplasias definitorias de SIDA Neoplasias no definitorias de SIDA El grado de inmunosupresión manifestado
CRITERIOS DE HORMONOSENSIBILIDAD. Dra. Nuria Ribelles. Hospital Universitario Virgen de la Victoria. Málaga.
 CRITERIOS DE HORMONOSENSIBILIDAD. Dra. Nuria Ribelles. Hospital Universitario Virgen de la Victoria. Málaga. Criterios generales Receptor Estrógeno Receptor Progesterona HER2 Biopsia de la metástasis Nuevos
CRITERIOS DE HORMONOSENSIBILIDAD. Dra. Nuria Ribelles. Hospital Universitario Virgen de la Victoria. Málaga. Criterios generales Receptor Estrógeno Receptor Progesterona HER2 Biopsia de la metástasis Nuevos
Estructura y dinámica del citoesqueleto
 Estructura y dinámica del citoesqueleto Dr. Julio Tapia Pineda Programa de Biología Celular y Molecular ICBM, Facultad de Medicina TERAPIA OCUPACIONAL Abril 2012 Compartimentalización de una célula eucarionte
Estructura y dinámica del citoesqueleto Dr. Julio Tapia Pineda Programa de Biología Celular y Molecular ICBM, Facultad de Medicina TERAPIA OCUPACIONAL Abril 2012 Compartimentalización de una célula eucarionte
CONICYT: Repositorio Institucional: Ficha de Iniciativa de CIT (Ciencia, Tecnología e Innovación) 1 FONDECYT-REGULAR - 2001-1010800
 1 FONDECYT-REGULAR - 2001-1010800") CONICYT: Repositorio Institucional: Ficha de Iniciativa de CIT (Ciencia, Tecnología e Innovación) 1 FONDECYT-REGULAR - 2001-1010800 CANCER DE MAMA: ESTUDIO EN MUJERES CHILENAS CON HISTORIA FAMILIAR PARA
CONICYT: Repositorio Institucional: Ficha de Iniciativa de CIT (Ciencia, Tecnología e Innovación) 1 FONDECYT-REGULAR - 2001-1010800 CANCER DE MAMA: ESTUDIO EN MUJERES CHILENAS CON HISTORIA FAMILIAR PARA
Marta Hergueta Redondo Dic. -2005
 Perfil de expresión genética en el carcinoma no microcítico de pulmón: utilidad de las biopsias endoscópicas, correlación con la histología y predicción pronóstica en pacientes operables. FIS 04/2641 Marta
Perfil de expresión genética en el carcinoma no microcítico de pulmón: utilidad de las biopsias endoscópicas, correlación con la histología y predicción pronóstica en pacientes operables. FIS 04/2641 Marta